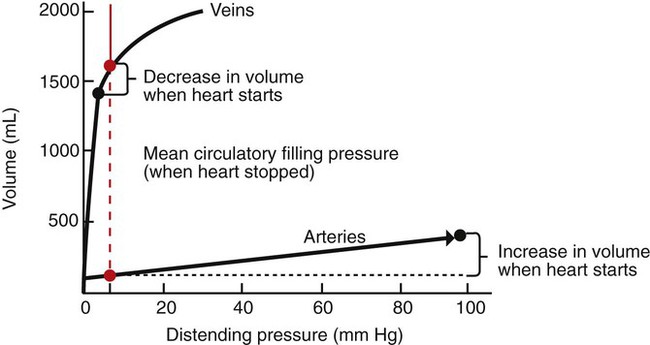
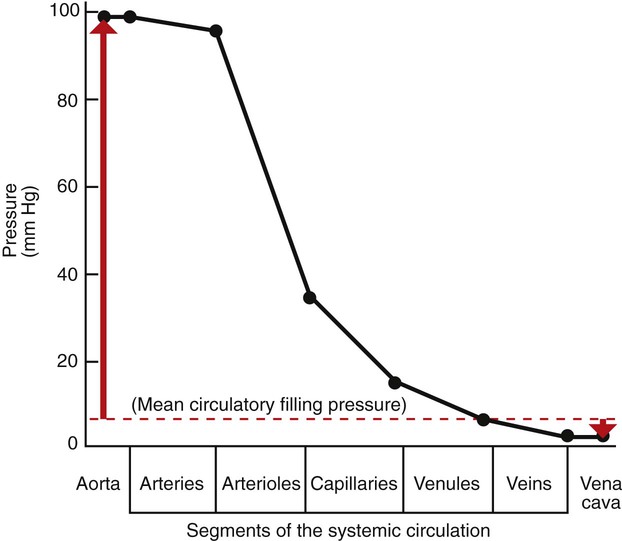
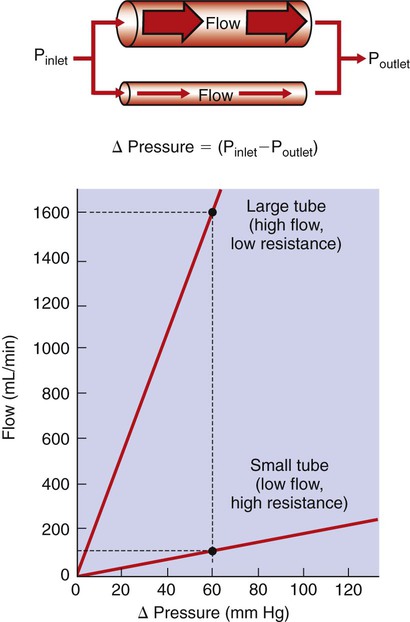
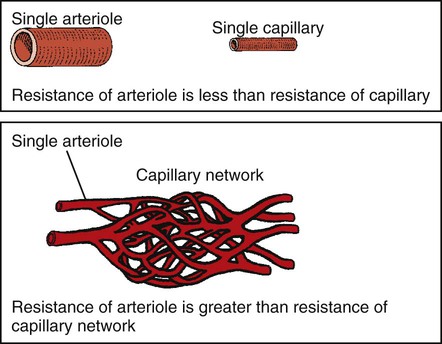
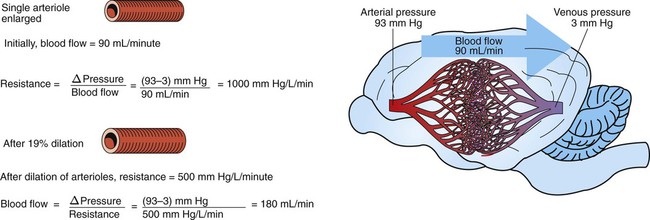
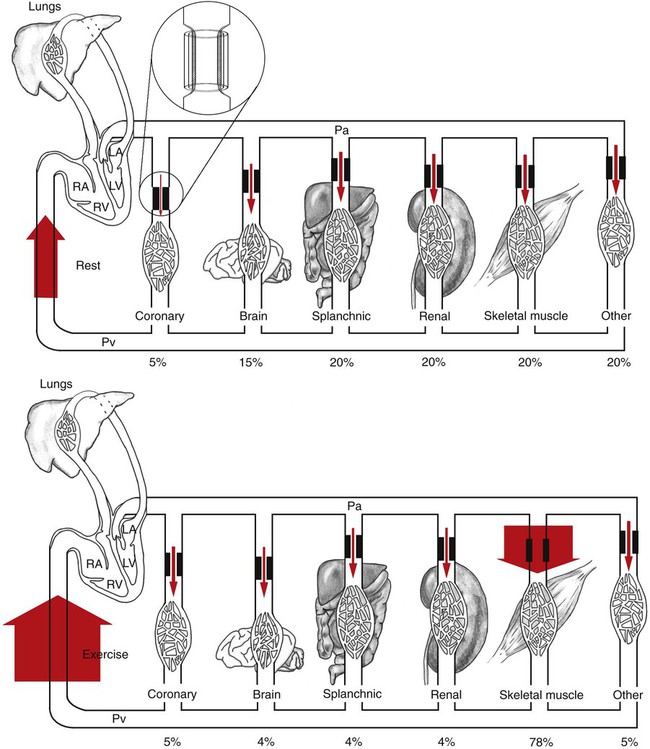
1. Blood pressure represents a potential energy that propels blood through the circulation. 2. Vascular resistance is defined as perfusion pressure divided by flow. 3. The net resistance of the systemic circulation is called the total peripheral resistance. 4. Arterial pressure is determined by the cardiac output and the total peripheral resistance. 5. Blood flow to each organ is determined by perfusion pressure and by the organ’s vascular resistance. 6. The pulmonary circulation offers much less resistance to blood flow than does the systemic circulation. 7. Arterial pressures are measured in terms of systolic, diastolic, and mean levels. 8. Pulse pressure increases when the stroke volume increases, heart rate decreases, aortic compliance decreases, or total peripheral resistance increases. Figure 22-1 shows the normal pressure profile in the systemic circulation. This figure portrays the pressures that would be measured if a miniature pressure gauge were inserted into the various vessels that blood passes through in its journey through the systemic circulation. The blood pressure is highest in the aorta (typically, mean aortic pressure is 98 mm Hg) and lowest in the venae cavae (typically, 3 mm Hg). The difference between these pressures (i.e., 95 mm Hg) constitutes the driving force for the movement of blood, by bulk flow, through the systemic circulation. As discussed in Chapter 18, such a pressure difference between the inlet and outlet of a tube (or system of tubes) is called perfusion pressure difference (or more commonly, just perfusion pressure). Consider what happens if the heart is restarted in an animal after the pressure has equalized everywhere at 7 mm Hg. With each heartbeat, the heart takes some blood out of the venae cavae and this volume of blood is transferred (via the pulmonary circulation) into the aorta. The volume of blood in the venae cavae decreases, so the venae cavae become less distended and vena caval pressure drops below 7 mm Hg. The volume of blood in the aorta increases, so the aorta becomes more distended and aortic pressure rises above 7 mm Hg. As illustrated in Figure 22-1, the vena caval pressure drops about 4 mm Hg (from 7 to 3 mm Hg), and the aortic pressure rises about 91 mm Hg (from 7 to 98 mm Hg). It is important to understand why the pressure decreases only a little in the venae cavae but increases so much in the aorta, even though the volume of blood removed from the venae cavae with each heartbeat is the same as the volume of blood added to the aorta. The reason is that the veins are much more compliant (distensible) than the arteries; one can add or remove blood from veins without changing the venous pressure very much, whereas the addition or removal of blood from arteries changes the arterial pressure a great deal. Compliance corresponds to the slope of a volume-versus-pressure graph. As illustrated in Figure 22-2, veins are about 20 times more compliant than arteries (over the range of pressures typically encountered in the circulation). Therefore, veins can accept or give up a large volume of blood without incurring much of a change in pressure. Veins readily expand or contract to accommodate the changes in blood volume that occur with fluid intake (e.g., drinking) or fluid loss (e.g., sweating). Veins thus function as the major blood volume reservoirs of the body. In contrast, arteries function as pressure reservoirs, providing the temporary storage site for the surge of pressure energy that is created with each cardiac ejection. Arteries are tough vessels, with low compliance. Therefore, arteries can accommodate a large increase in pressure during a cardiac ejection and then sustain the pressure high enough between cardiac ejections to provide a continuous flow of blood through the systemic circulation. Where Δ Pressure is perfusion pressure difference, or simply perfusion pressure (i.e., the pressure at the tube inlet minus the pressure at its outlet). Figure 22-3 presents these concepts in pictorial and graphic form. The dashed lines in this figure indicate that a perfusion pressure of 60 mm Hg causes a flow of 1600 milliliters per minute (mL/min) through the large tube. Thus the resistance of the large tube is 37.5 mm Hg/liter per minute (L/min). The same perfusion pressure (60 mm Hg) causes a flow of only 100 mL/min through the small tube. The resistance of the small tube is therefore 600 mm Hg/L/min. The resistance of the small tube is 16 times greater than the resistance of the large tube. As already described, the arterioles are the segment of the systemic circulation with the highest resistance to blood flow (see Figure 22-1). It may seem paradoxical that the arterioles are the site of highest resistance when the capillaries are smaller vessels. After all, Poiseuille’s law and Figure 22-3 emphasize that a smaller tube has a much higher resistance than a larger tube. The resolution of this paradox is presented in Figure 22-4. It is true that each capillary has a smaller radius and therefore a greater resistance than each arteriole. However, each arteriole in the body distributes blood to many capillaries, and the net resistance of all those capillaries is less than the resistance of the single arteriole that delivers blood to them. It is only because each arteriole delivers blood to so many capillaries that the net resistance of the capillaries is less than the resistance of the arteriole. Figure 22-5 illustrates that a small change in the radius of arterioles in an organ brings about a large change in resistance and therefore in blood flow. In this example the arterial pressure is 93 mm Hg and the venous pressure is 3 mm Hg, so the perfusion pressure is 90 mm Hg. The brain blood flow is initially observed to be 90 mL/min. Based on the mathematical definition of resistance, the resistance of the brain blood vessels is 1000 mm Hg/L/min. Most of this resistance is provided by the brain arterioles. Next, consider the consequence of a slight vasodilation, such that the radius of the arterioles increases by 19% (e.g., from a radius of 1.00 to a radius of 1.19). Recall from Poiseuille’s law that the resistance varies inversely as the fourth power of the radius. Because 1.194 equals 2.00, a 19% increase in radius cuts the resistance in half. Decreasing the brain’s resistance by half (to 500 mm Hg/L/min) would double the brain blood flow (to 180 mL/min). The response to vigorous exercise provides a third application of the concept that the mean aortic blood pressure is determined by the cardiac output and TPR. As in hemorrhage, exercise causes the cardiac output and TPR to change in opposite directions. In exercise, however, the cardiac output is elevated, and TPR is decreased. TPR decreases because the arterioles in the exercising skeletal muscle dilate, which increases skeletal muscle blood flow. During vigorous exercise, TPR decreases to about one-fourth of its resting value. The cardiac output increases about fourfold. The result is that the aortic pressure is negligibly changed. Figure 22-6 depicts the cardiovascular adjustments to vigorous exercise.
The Systemic and Pulmonary Circulations
Blood Pressure Represents a Potential Energy That Propels Blood Through the Circulation
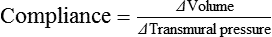
Vascular Resistance Is Defined as Perfusion Pressure Divided by Flow
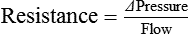
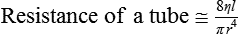
Arterial Pressure Is Determined by the Cardiac Output and the Total Peripheral Resistance
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The Systemic and Pulmonary Circulations

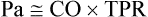
Only gold members can continue reading. Log In or Register to continue
