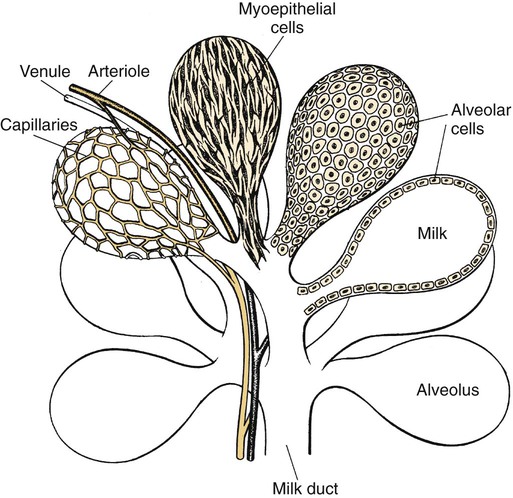
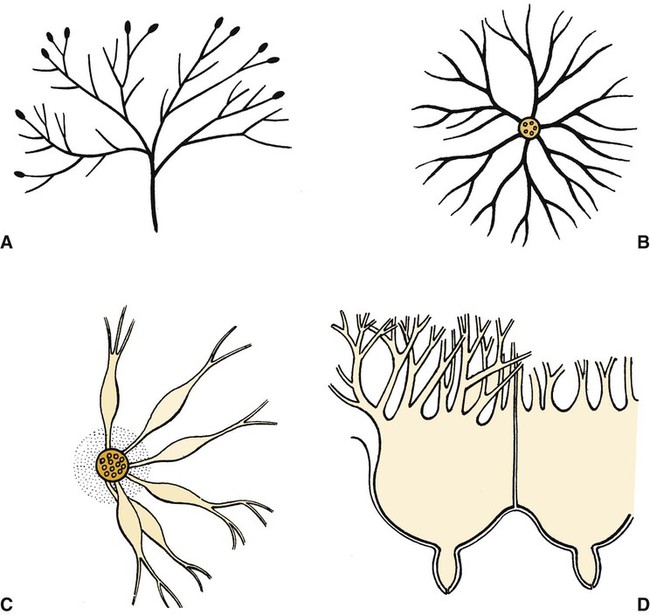
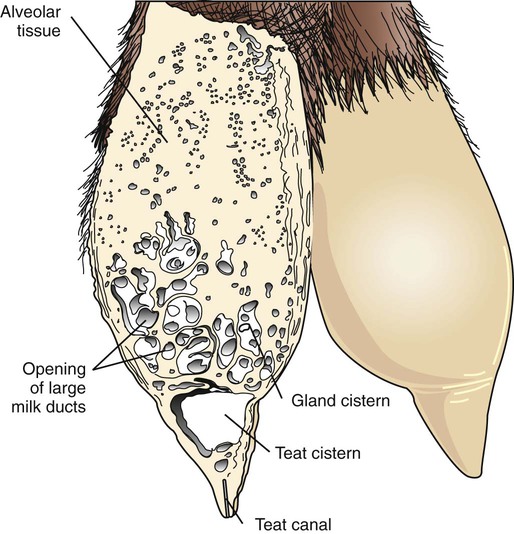
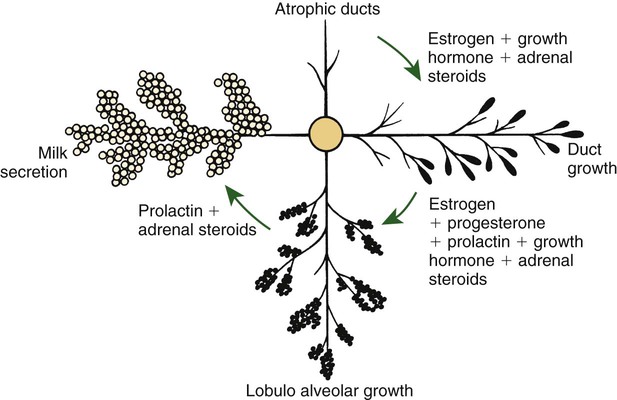
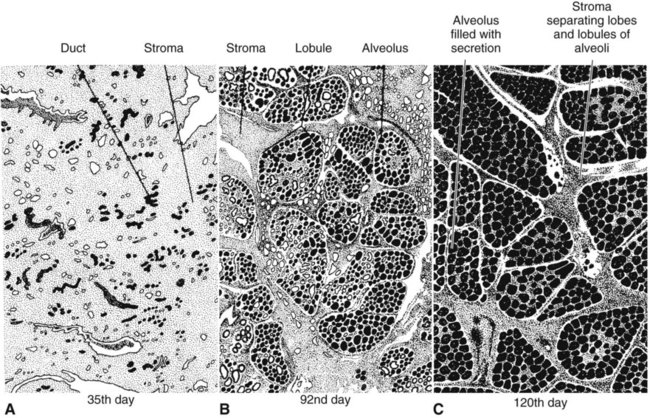
Anatomical aspects of the mammary gland 1. The milk-secreting cells of the mammary gland develop through the proliferation of epithelium into hollow structures called alveoli. 2. Most of the milk that accumulates before suckling or milking is stored in the alveoli, even though animals have enlarged milk-storage areas called cisterns. 3. A suspensory system involving the udder of the cow allows the animal to carry a large amount of milk. 1. Initial development of the mammary gland is programmed by embryonic mesenchyme. 2. Proliferation of the mammary duct system begins at puberty, with ducts under the control of estrogens, growth hormone, and adrenal steroids, and alveoli under the control of progesterone and prolactin. 1. Prepartum milk secretion (without removal) results in the formation of colostrum. 2. The ingestion of colostrum is important because of the passive immunity it confers through the presence of high concentrations of immunoglobulins. 3. The time immunoglobulins can be absorbed through the neonatal gut is limited to the first 24 to 36 hours of life. 4. Lipids (particularly vitamin A) and proteins (caseins and albumins) are high in concentration in colostrum; carbohydrates (lactose) are low. 1. Prolactin, inhibited by dopamine and stimulated by vasoactive intestinal peptide, is the most important hormone involved in the process of milk synthesis, or lactogenesis; growth hormone is also important for lactogenesis. 2. The release of fat into milk from the alveolar cell involves constriction of the plasma membrane around the fat droplet; fats are dispersed in milk in droplet form. 3. Milk proteins and lactose are released from alveolar cells by the process of exocytosis. 1. Efficient milk removal requires the release of oxytocin, which causes contraction of muscle cells that surround the alveoli (myoepithelial cells), and movement of milk into the ducts and cisterns. 1. Carbohydrate stores are good in neonates born as singles or twins, whereas carbohydrate stores are low in neonates born in litters; consequently the former can stand a longer interval to first suckling than can the latter. 1. Fats are the most important energy source in milk. 2. Lactose, composed of glucose and galactose, is the main carbohydrate of mammalian milk. 3. The main proteins in milk are called caseins and are found in curd. 1. Milk production peaks at 1 month postpartum in dairy cattle, followed by a slow decline in production; milking usually stops at 305 days of lactation so that the animal can prepare the mammary gland for the next lactation. 2. Lactation can be induced by hormone administration (estrogen and progesterone) and enhanced by growth hormone and increased photoperiod exposure. Diseases associated with the mammary gland 1. The main diseases that affect the mammary gland directly are mastitis (prevalent in dairy cattle and dogs) and neoplasia (prevalent in intact dogs and cats). 2. The main conditions that involve the mammary gland indirectly are passive transfer of red blood cell agglutinating antibodies by the ingestion of colostrum (mare, queen) and hypocalcemia caused by the transient drain of calcium that occurs with initiation of lactation (dairy cattle) or during the perinatal period (dog). The parenchyma, or milk-secreting cells, of the mammary gland develops through the proliferation of epithelial cells that arise from the primary mammary cord. The epithelial cells eventually form hollow, circular structures called alveoli, which are the fundamental milk-secreting units of the mammary gland (Figure 39-1). In concert with this development, an enlarged area of epithelium, the nipple, which is the external connection to the internal milk-secreting system, develops on the surface. In males, although nipples often develop, the underlying primary mammary cord does not develop into substantial glandular tissue. Duct systems connect alveoli with the nipple, or teat, enabling milk to pass from the area of formation to the area of delivery (nipple). The ducts may come together so that there is only one final duct per gland, which has one opening through the nipple, or teat, such as occurs in cattle, goats, and sheep. Two main ducts and associated openings occur in the mare and sow, whereas the cat and dog can have 10 or more openings in the nipple, with each opening representing separate glands (Figure 39-2). Both the cow and the doe (goat) have specialized areas for holding milk, called cisterns, which are located in the ventral part of the gland and into which all main ducts empty (Figure 39-3). This has enabled the cow, for example, to synthesize and store larger amounts of milk than would otherwise be possible. Despite this adaptation, it is important to realize that a majority of the milk present at the time of milking is stored in the duct system of the mammary glands. Development of the mammary gland in post-fetal life usually starts in concert with puberty. Cyclical ovarian activity results in the production of estrogen and progesterone. Estrogen, with growth hormone and adrenal steroids, is responsible for proliferation of the duct system. The development of alveoli from the terminal ends of the ducts requires the addition of progesterone and prolactin (Figure 39-4). Although the development of the mammary gland begins with the onset of puberty, the gland remains relatively undeveloped until the occurrence of pregnancy. In most domestic animals, udder development usually becomes evident by the middle of gestation; the secretion of milk often begins during the latter part of gestation (mainly from increasing prolactin secretion) and results in the formation of colostrum, as discussed later. By the end of pregnancy, the mammary gland has been transformed from a structure involving mostly stromal (connective tissue) elements to a structure that is filled with alveolar cells that are actively synthesizing and secreting milk. Groups of adjacent alveoli form lobules that further combine into larger structures called lobes. Connective tissue bands delineate the lobules and the lobes (Figure 39-5). Colostrum is a rich source of nutrients, especially vitamin A, in addition to immunoglobulins. Placental transfer of vitamin A is limited in domestic animals, with calves and piglets being particularly low in vitamin A at birth. This deficiency is corrected by the ingestion of colostrum. Lipids and proteins, including caseins and albumins, are also present in relatively high concentration in colostrum. One exception is lactose; its synthesis is significantly inhibited by progesterone until about the time of delivery. Nevertheless, at the moment of delivery, the newborn’s milk supply is nutritive (high protein, fat, and vitamin A content) and protective (immunoglobulins) (Table 39-1). TABLE 39-1 Amounts of Selected Components of Bovine Colostrum as Percentage of Level in Normal Milk
The Mammary Gland
Anatomical Aspects of the Mammary Gland
The Milk-Secreting Cells of the Mammary Gland Develop Through the Proliferation of Epithelium into Hollow Structures Called Alveoli
Most of the Milk That Accumulates Before Suckling or Milking Is Stored in the Alveoli, Even Though Animals Have Enlarged Milk-Storage Areas Called Cisterns
Control of Mammogenesis
Initial Development of the Mammary Gland Is Programmed by Embryonic Mesenchyme
Proliferation of the Mammary Duct System Begins at Puberty, with Ducts Under the Control of Estrogens, Growth Hormone, and Adrenal Steroids, and Alveoli Under the Control of Progesterone and Prolactin
Colostrum
Prepartum Milk Secretion (Without Removal) Results in the Formation of Colostrum
Lipids (Particularly Vitamin A) and Proteins (Caseins and Albumins) Are High in Concentration in Colostrum; Carbohydrates (Lactose) Are Low
Constituent
Day(s) After Parturition
0
3
4
Dry matter
220
100
100
Lactose
45
90
100
Lipids
150
90
100
Minerals
120
100
100
Proteins
Casein
210
110
110
Albumin
500
120
105
Globulin
3500
300
200
Vitamins
A
600
120
100
Carotene
1200
250
125
E
500
200
125
Thiamine
150
150
150
Riboflavin
320
130
110
Pantothenic acid
45
110
105 ![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The Mammary Gland
Only gold members can continue reading. Log In or Register to continue
