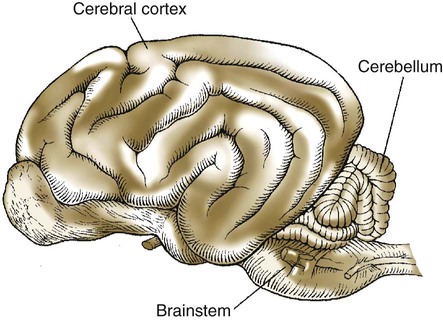
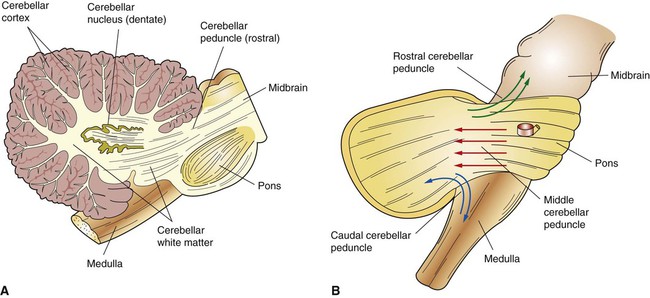
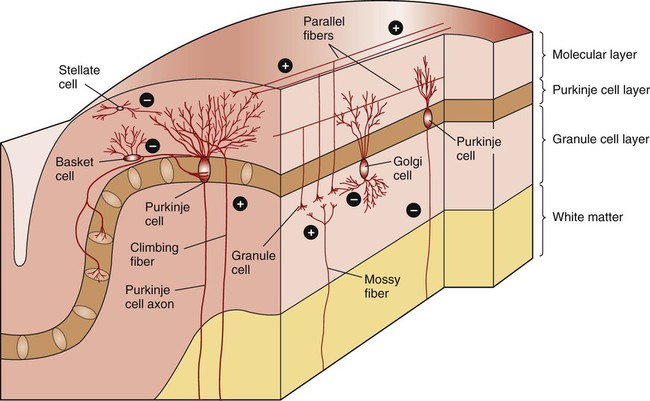
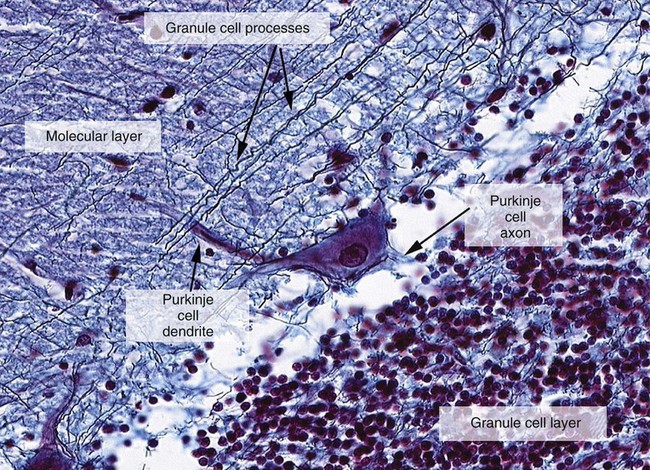
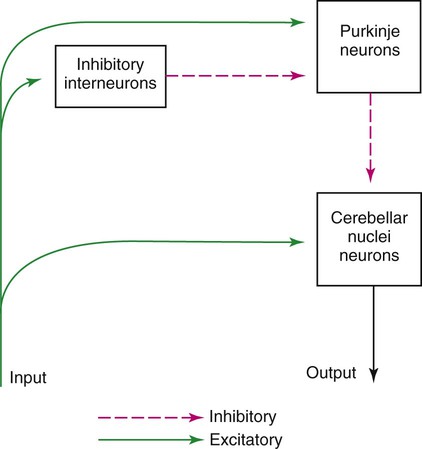
1. The cerebellum constantly compares the intended movement with the actual movement and makes appropriate adjustments. 2. Cerebellar histology and phylogeny give clues to cerebellar function. 3. The vestibulocerebellum helps coordinate balance and eye movements. 4. The spinocerebellum helps coordinate muscle tone as well as limb movement. 5. The cerebrocerebellum helps with planning coordinated, properly timed movement sequences. 6. The cerebellum plays a role in motor learning. 7. Cerebellar disease causes abnormalities of movement and further illuminates cerebellar function. The cerebellum (Latin, “little brain”) is caudal to the cerebral cortex and dorsal to the brainstem (Figure 12-1). Although it constitutes only about 10% of the gross brain volume because of its highly folded structure, the cerebellum contains more than half of all CNS neurons. The outer layer of cerebellar gray matter, the cerebellar cortex, has a highly regular, three-layered, histological appearance, which suggests that all cerebellar regions may perform a common underlying task. Like the cerebral cortex, the particular inputs to a given region of cerebellar cortex, and the particular output targets that it influences, in large part account for the functional differences between cerebellar regions. In addition to the cerebellar cortex, and the cerebellar white matter axons entering and leaving the cortex, a group of deep cerebellar nuclei is embedded within the cerebellar white matter (Figure 12-2). The cells of these nuclei are a principal origin of the axons leaving the cerebellum. Two large pairs of white matter stalks, the rostral and middle cerebellar peduncles, respectively carry axons out from and into the cerebellum. A third, smaller pair of cerebellar peduncles, the caudal cerebellar peduncles, carry axons both into and out from the cerebellum. The cerebellum is not necessary for the initiation of movement. Muscle strength remains largely intact with complete destruction of the cerebellum. However, the cerebellum plays a crucial role in the timing and coordination of movement initiated by the parts of the motor system hierarchy discussed in Chapter 10. It does so by adjusting and modulating the output of the motor cortices, corticospinal tract, descending brainstem motor pathways, and spinal cord. Lesions of the cerebellum lead to major clinical deficits in the precision and grace with which movement is accomplished. The cortex throughout the cerebellum is quite uniform and consists of three layers and only five types of neurons: stellate, basket, Golgi, granule, and Purkinje cells (Figure 12-3). The outermost layer is the molecular layer and consists primarily of granule cell axons, known as parallel fibers (Figure 12-4); dendrites of neurons located in deeper layers; and scattered inhibitory interneurons, the stellate and basket cells. The middle Purkinje cell layer of cerebellar cortex consists of the large cell bodies of Purkinje neurons, which have a flat but extremely expansive dendritic field that extends into the molecular layer (see Figures 12-3 and 12-4). This dendritic field is oriented at right angles to the parallel fibers. Therefore, a Purkinje cell is contacted by an expansive array of parallel fiber axons of granule cells, and an individual parallel fiber contacts the dendrites of many Purkinje cells. The stellate and basket cell inhibitory interneurons, noted above, can act to refine, or prune, this extensive spatial pattern of Purkinje cell activation by parallel fibers. The innermost granule cell layer of cerebellar cortex contains the vast number of granule cell somas that give rise to the parallel fibers (see Figures 12-3 and 12-4). This layer also contains occasional Golgi cell bodies. These are inhibitory interneurons that can regulate the overall level of excitation of the Purkinje cells by the granule cell parallel fibers. Axons of the Purkinje neurons go to the deep cerebellar nuclei, located outside of the cerebellar cortex, embedded in the cerebellar white matter (see Figure 12-2). The Purkinje cells are the only output neurons of the cerebellar cortex and are all inhibitory. They can inhibit the spontaneously active neurons of the deep cerebellar nuclei, whose axons leave the cerebellum. This selective inhibition represents a sensitive temporal refinement of cerebellar processing that supplements the spatial refinement, and the excitation level control, noted above. The cerebellar output neurons participate in regulating the activity of brainstem motor pathways and motor cortices involved in the execution and planning of movement. The two primary groups of input axons to the cerebellum are the mossy fiber and climbing fiber axons (see Figure 12-3). Both are excitatory; they cause excitatory postsynaptic potentials (EPSPs) within the cerebellar cortex and, through collateral axons, within the deep cerebellar nuclei (Figure 12-5). The mossy and climbing fibers collectively carry information from components of the motor system hierarchy and from peripheral sensory receptors regarding the planning, initiation, and execution of the movement. The shorter input/output circuit of the cerebellum consists of the climbing and mossy fiber stimulation to the deep cerebellar nuclei, whose output in turn leaves the cerebellum to modify components of the motor system hierarchy. However, the output of the deep cerebellar nuclei is itself modified by inhibition from Purkinje cell axons that originate in cerebellar cortex. The Purkinje cell inhibition of deep cerebellar nuclei is based on the cerebellar cortex’s own integration of mossy and climbing fiber inputs. In other words, the same information coming into the cerebellum that drives the cerebellar nuclei is also processed by cerebellar cortex, whose resulting Purkinje cell output refines or “sculpts” the output of the cerebellar nuclei that project to components of the motor system. Within the cerebellar cortex, inhibitory interneurons help to refine or “sculpt” the Purkinje cell output of cerebellar cortex.
The Cerebellum
Cerebellar Histology and Phylogeny Give Clues to Cerebellar Function
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The Cerebellum
Only gold members can continue reading. Log In or Register to continue
