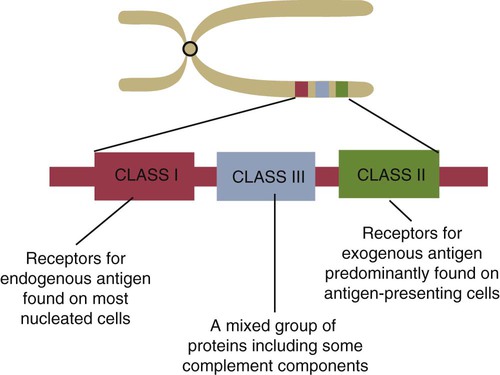
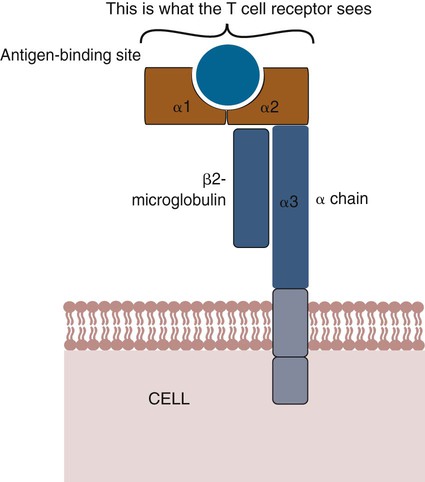
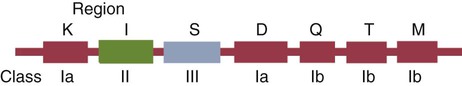
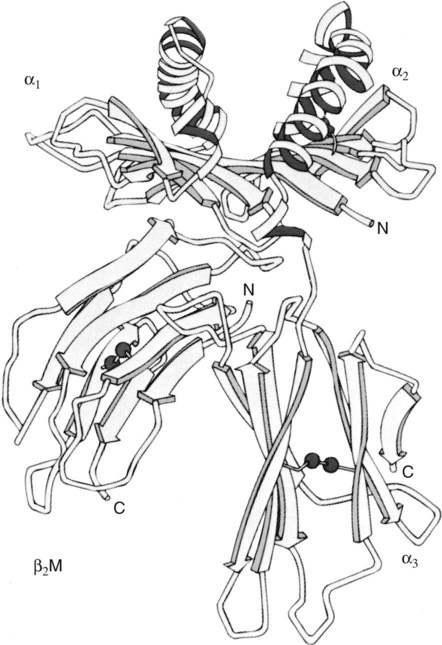
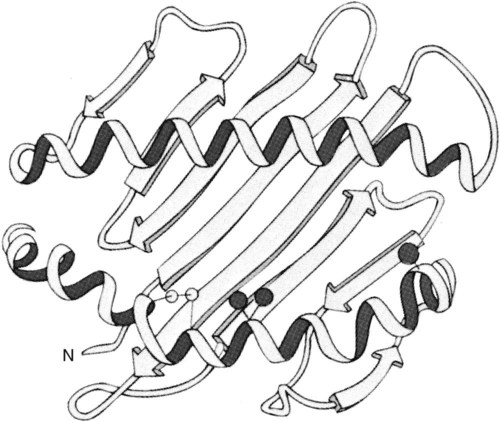
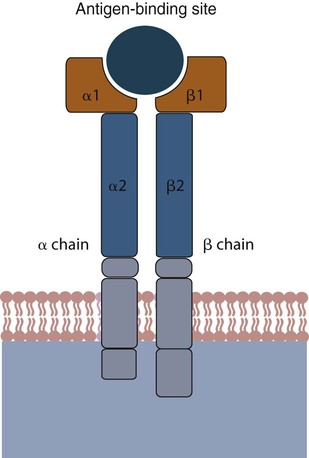
• Antigen-presenting cells use receptors called MHC molecules to bind and present antigens. • MHC molecules are encoded by genes located within the major histocompatibility complex. • The classical MHC molecules are highly polymorphic. That is, they show an enormous variety of inherited structural variations that permit each individual animal to respond to a different set of antigens. • Class I MHC molecules are found on all nucleated cells. Their function is to present endogenous antigens to CD8+ T cells. • Class II MHC molecules are largely restricted to professional antigen-presenting cells such as dendritic cells, macrophages, and B cells. Their function is to present exogenous antigens to CD4+ T cells. • The class III region of the MHC contains a mixture of genes, some of which encode complement components. In order to trigger an adaptive immune response, antigen molecules must be broken up inside cells, and the fragments generated must then be bound to appropriate antigen-presenting receptors (Figure 11-1). These antigen-presenting receptors are glycoproteins encoded by genes located in a large gene cluster called the major histocompatibility complex (MHC). The receptors are therefore called MHC molecules. Antigen fragments can trigger an immune response only after they have bound to MHC molecules, and these antigen-MHC complexes have bound to T cell antigen receptors. Since the MHC molecules serve as specific antigen receptors, MHC genes determine which antigens can trigger adaptive immunity. Thus the MHC can be considered an organized cluster of genes that control antigen presentation and so determine an animal’s susceptibility to infectious or autoimmune diseases. All vertebrates, from cartilaginous fish to mammals, possess cell-surface proteins coded for by genes clustered within an MHC. Each MHC has a fairly consistent structure consisting of about 200 expressed genes distributed over about 3 mb of DNA and divided into three regions containing different classes of MHC gene loci (I, II, and III) (Figure 11-2). Classical class I loci code for the MHC molecules expressed on most nucleated cells. Class I genes can be subdivided into those that are highly polymorphic (class Ia genes) and those that show very little polymorphism (class Ib, Ic, or Id genes). (Polymorphism refers to structural variations between proteins.) Class Id genes are located outside the MHC on a different chromosome. Genes in class II loci, on the other hand, encode polymorphic MHC molecules mainly restricted to professional antigen-presenting cells (dendritic cells, macrophages, and B cells) (Table 11-1). Genes in the class III region code for a diverse mixture of proteins, many of which are linked to innate immunity, such as complement proteins. Although each MHC contains all three gene regions, their gene content, and arrangement vary between species. Table 11-1 Comparison of MHC Class I and Class I Structure Class Ia molecules consist of two linked glycoprotein chains. An α chain (45 kDa) is associated with a much smaller chain called β2-microglobulin (β2M) (12 kDa). The α chain is inserted in the cell membrane (Figure 11-3). It consists of five domains: three extracellular domains called α1, α2, and α3, each about 100 amino acids long; a transmembrane domain; and a cytoplasmic domain. The antigen-binding site on these molecules is formed by the α1 and α2 domains. β2M consists of a single domain and serves to stabilize the structure. The size of the MHC class I region varies among mammals. Humans and rodents have the largest, and pigs have the smallest. The chicken class I region is very much smaller than in mammals (Chapter 40). The MHC class I region has a common framework of non-MHC genes, and size differences are mainly due to variations in the size and number of these framework genes. The number of class Ia gene loci varies between mammals. For example, rats have more than 60, mice have about 30, humans have 20, cattle have 13 to 15, and pigs have 11. Not all these genes are functional. For example, in mice, only two or three class I genes are expressed. The remainder are pseudogenes (defective genes that cannot be expressed). In humans the functional polymorphic genes are called A, B, and C. In mice they are called K and D (and in some strains, L) (Figure 11-4). In other species they are usually numbered. The α1 and α2 domains of MHC class I molecules fold together to form an open-ended groove (Figure 11-5). A flat β sheet forms the floor of this groove, and its walls are formed by two α helices (Figure 11-6). This groove binds antigenic peptides, 8 to 10 amino acids in size. The variable regions located along the walls of this groove determine its shape. The shape of the groove in turn determines which peptides can be bound and thus trigger immune responses. Class Ib molecules show reduced expression and tissue distribution compared with class Ia molecules but are part of the MHC complex. They have limited polymorphism and their genes probably originated from class Ia precursors by gene duplication. For example, the class Ib genes in mice are found in three loci called Q, T, and M (see Figure 11-4). They code for proteins on the surface of regulatory and immature lymphocytes and on hematopoietic cells. These also consist of a membrane-bound α chain associated with β2-microglobulin, so their overall shape and antigen-binding groove are similar to those in MHC class Ia molecules. Since they are not polymorphic, MHC class Ib molecules bind a limited range of ligands. Thus they are receptors for commonly encountered, microbial pathogen-associated molecular patterns (PAMPs). Class Ic genes have limited polymorphism and are found within the MHC but probably originated before the radiation of the placental mammals. Their products include MICA and MICB, specialized proteins that are involved in signaling to natural killer (NK) cells but do not bind antigenic peptides (Chapter 19). Class Id genes are nonpolymorphic class I–related genes not located on the MHC chromosome. Many of their products contribute to innate immunity since they bind PAMPs. For example, CD1 molecules bind bacterial lipids (Chapter 19). FcRn is a class Id MHC molecule that serves as an antibody (Fc) receptor on epithelial cells. It is expressed on mammary gland epithelium and on the enterocytes of newborn mammals (Chapter 21). Mammals differ in their expression of MHC class II molecules. In rodents, they are restricted to the professional antigen-presenting cells (dendritic cells, macrophages, and B cells) but can be induced on T cells, keratinocytes, and vascular endothelial cells. Resting mouse T cells do not express MHC class II molecules, but in pigs, dogs, cats, mink, and horses, the MHC class II molecules are constitutively expressed on nearly all resting adult T cells. In cattle, most MHC class II molecules are expressed only on B cells and activated T cells. In pigs, resting T cells express MHC class II molecules at about the same level as macrophages. In humans and pigs, MHC class II molecules are expressed on renal vascular endothelium and glomeruli—a fact of significance in kidney graft rejection. The expression of class II molecules is enhanced in rapidly dividing cells and in cells treated with interferon-γ (IFN-γ) (Chapter 14). MHC class II molecules consist of two chains called α and β. Each chain has two extracellular domains (one constant and one variable), a connecting peptide, a transmembrane domain, and a cytoplasmic domain (Figure 11-7). A third chain, called the Ii or γ chain, is associated with the assembly of class II molecules within cells and was discussed in Chapter 10.
The Major Histocompatibility Complex
Major Histocompatibility Complex
CLASS I
CLASS II
Loci include
Typically A, B, and C
DP, DQ, and DR,
Distribution
Most nucleated cells
B cells, macrophages, and dendritic cells
Function
Present antigen to cytotoxic T cells
Present antigen to T helper cells
Result
T-cell–mediated toxicity
T-cell–mediated help
MHC Class Ia Molecules
Structure
Gene Arrangement
Polymorphism
Nonpolymorphic MHC Class I Molecules
MHC Class II Molecules
Structure
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The Major Histocompatibility Complex

Only gold members can continue reading. Log In or Register to continue
