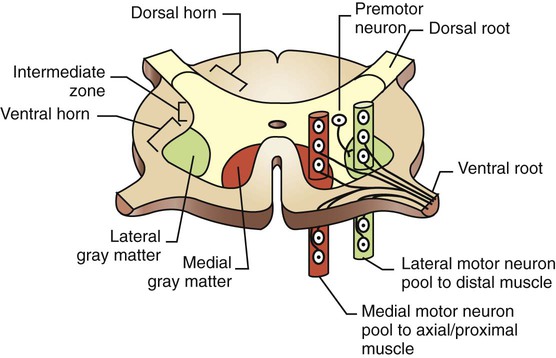
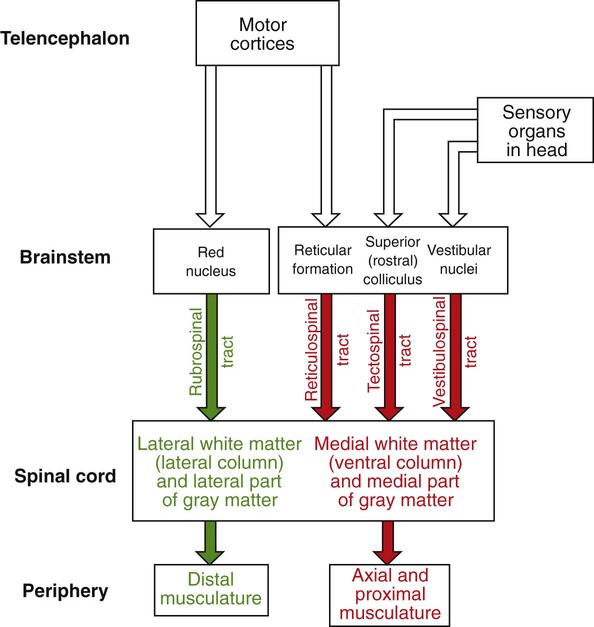
1. The central nervous system structures that control movement have a hierarchical organization. 2. The spinal cord is the most caudal and simplest level of the movement control hierarchy. 3. Brainstem upper motor neuron pathways are the source of all descending motor system input to the spinal cord, except for one other major pathway. 4. Medial and lateral descending brainstem motor pathways respectively control proximal muscles of posture and more distal muscles of skilled movement. 5. The reticulospinal and vestibulospinal tracts are medial brainstem motor pathways important for keeping the body upright against the pull of gravity. 6. The rubrospinal tract is a lateral brainstem motor pathway that can control distal limb musculature associated with skilled movement. 7. The corticospinal (pyramidal) tract is a direct projection from cerebral cortex to spinal cord responsible for the most skilled voluntary movements of mammals. 8. The corticospinal tract has a massive lateral component controlling the distal musculature and a minor medial component controlling the axial and proximal musculature. 9. The motor cortices of the frontal lobe, the highest level of the motor control hierarchy, consist of three different functional regions. 10. Corticospinal tract co-activation of both alpha (α) and gamma (γ) lower motor neurons may help with small automatic corrections of voluntary movements. 11. The motor system shares some organizational principles with sensory systems. 12. The basal ganglia and cerebellum modulate the activity of motor system components for the respective selection and adjustment of movement. Unlike the sensory systems, most of which transform physical energy into neural information, the motor system transforms neural information into physical energy. All movement is the result of the contraction of varying numbers of extrafusal skeletal muscle fibers within varying numbers of motor units (see Figure 6-8). These extrafusal muscle fibers do not contract until commanded to do so by the alpha (α) lower motor neuron. The α motor neuron, in turn, does not send such an action potential command until signaled to do so by descending upper motor neurons (see Figure 9-1) or from incoming sensory neurons (or interneurons) in a reflex arc. Another organizational principle of the neural control of movement is that it consists of a hierarchy. Generally, simpler movements or movement patterns are organized by more caudal parts of the CNS (Figure 10-1, bottom portions), and more complex and skilled patterns are organized by progressively more rostral regions (see Figure 10-1, top portions). The spinal cord contains the lower motor neurons that synapse upon the muscles of the trunk and limbs (see Figure 10-1). As noted in Chapter 6, an α lower motor neuron innervates several extrafusal muscle fibers of a single muscle, forming a motor unit (see Figure 6-8). The neuronal cell bodies of the motor units of a given muscle are clustered into a motor neuron pool located in the ventral horn of spinal cord gray matter. The motor neuron pool of a muscle has a cigar-shaped, longitudinal organization in the cord, often extending rostrocaudally over a few spinal cord segments (e.g., L1-L3; Figure 10-2). These motor neuron pools have a somatotopic organization in the ventral horn; that is, their relative position in the CNS corresponds to the relative body position of the muscles that their neurons innervate. In other words, motor neuron pools whose neurons innervate distal muscles of the limbs tend to be located in more lateral parts of the ventral horn, whereas motor neuron pools associated with axial and proximal musculature tend to be located more medially within the ventral horn. Lower motor neurons projecting out to muscles are often synaptically activated by premotor neurons whose cell bodies are usually located in the intermediate zone of spinal cord gray matter (see Figure 10-2). Activating a premotor neuron in the lateral part of the intermediate zone on one side of the body will generally activate a modest number of α motor neurons, in the lateral part of the ventral horn, on the same side of the body. This in turn will result in the activation of a modest number of distal limb muscles that would generally be used for skilled, voluntary movement. Premotor neuron activation in the medial part of the intermediate zone on one side of the body will generally activate a larger number of α motor neurons, in the medial part of the ventral horn, often on both sides of the body and often over more than one spinal cord segment. This in turn will result in the extensive activation of axial or proximal antigravity muscles on both sides of the body. Such a complement of muscles would be required for the involuntary stabilization or adjustment of posture. It can therefore be seen that more lateral parts of the spinal cord gray matter are involved in control of the distal limb musculature of skilled voluntary movement, whereas more medial parts are associated with the axial and proximal musculature of postural control. The simplest type of motor behavior, the spinal segmental reflex (e.g., the knee jerk reflex; see Chapter 7), can be organized at the level of the spinal cord, without significant control from more rostral divisions of the CNS (e.g., the brain). However, although control from the brain may not be necessary for such behaviors, these simple reflexes can often still be influenced by brain input. Furthermore, under different circumstances, the same spinal premotor and α motor neurons that participate in a simple spinal reflex could be activated by the brain to participate in an elegant and skilled sequence of movement. Four major axon tracts originate in the brainstem and descend to the spinal cord to influence spinal lower motor neurons: the vestibulospinal tract, the reticulospinal tract, the tectospinal tract, and the rubrospinal tract (Figure 10-3). Collectively, the first three are involved in the involuntary maintenance and adjustment of posture and in reflex orientation of the head. Therefore they are principally involved in the control of axial and proximal musculature. The rubrospinal tract is mainly involved in control of distal limb musculature of the type that mediates voluntary skilled movements. These four tracts (often along with components of the basal ganglia and cerebellum) are sometimes referred to as the extrapyramidal motor system. This is in contrast to the pyramidal motor system that originates in the cerebral cortex, the other major descending motor pathway to the spinal cord, as discussed later. Because the term extrapyramidal can encompass such a diverse group of structures, and because it is often applied inconsistently, it is being used less frequently. The four tracts from brainstem to spinal cord are collectively referred to here as the descending brainstem motor pathways. The brainstem, like the spinal cord, contains lower motor neurons that can synaptically activate skeletal muscles, in this case the face and head muscles (see Figure 10-1). The cell bodies of these α motor neurons reside in various cranial nerve nuclei (e.g., facial motor, hypoglossal, oculomotor). The brainstem also receives direct input from sensory organs in the face and head (e.g., eye, vestibular apparatus). Therefore, as in the spinal cord, some fairly simple segmental reflexes can be organized at the brainstem level without the necessity of significant control from other levels of the motor system. Because the brainstem also contains the descending motor pathways to the spinal cord previously noted, however, the brainstem also provides a means by which input from sensory organs in the face and head can reach and control lower motor neurons of the spinal cord that operate muscles of the trunk and limbs (see Figure 10-3). Some of the descending brainstem motor pathways also provide a means by which more rostral regions of the motor system (e.g., motor cortex) can indirectly influence spinal lower motor neurons. The descending brainstem motor pathways to the spinal cord can be divided into a medial group and a lateral pathway. The vestibulospinal, reticulospinal, and tectospinal tracts constitute the medial brainstem motor pathways, whereas the rubrospinal tract represents the lateral brainstem motor pathway (see Figure 10-3). The groupings are generally based on the relative position of these tracts within the spinal cord white matter. The axons of the tracts that represent the medial brainstem motor pathways (vestibulospinal, reticulospinal, tectospinal) will mainly travel in more medial regions of the spinal cord white matter (e.g., ventral column) and will synapse within more medial regions of the spinal cord gray matter. Those medial regions of the spinal gray matter contain medial premotor neurons and medial α motor neurons that control the axial and proximal extensor musculature primarily involved in involuntary maintenance and adjustment of posture. Axons of the lateral brainstem motor pathway (rubrospinal) run in a more lateral region of the spinal white matter (lateral column) and synapse in the more lateral spinal gray matter. The premotor and α motor neurons of this region principally control the distal flexor musculature involved in voluntary skilled movement. The reticulospinal tract originates from cell bodies in the reticular formation of the brainstem (see Figure 10-3). This is a netlike complex of many small clusters of cell bodies (nuclei) and loosely organized axonal projections, located near the midline. Once thought to be a diffuse and fairly nonspecific system, the reticular formation is now known to contain a number of functionally specific nuclei. In addition to being the origin of a medial descending brainstem motor pathway to the spinal cord, ascending projections of the reticular formation play an important role in modulating consciousness, arousal, and attention. The reticular formation receives a vast array of sensory information and plays an important role in pain perception, respiration, and circulatory function. Axons of the reticulospinal tract synapse within medial regions of the spinal cord gray matter that primarily control the axial and proximal extensor musculature (see Figure 10-3). Collectively, the tract projects to virtually all rostrocaudal levels of the cord. Portions of the reticulospinal tract that originate from cells in reticular nuclei of the pons tend to have an excitatory effect on lower motor neurons to the antigravity muscles. Portions of the tract coming from the reticular nuclei of the medulla tend to have an inhibitory effect on the lower motor neurons to antigravity muscles. These opposing portions of the reticulospinal tract interact to regulate antigravity muscle tone. Influences from other regions of the brainstem, the cerebellum, and the spinal cord endow the pontine reticular nuclei with a high level of spontaneous activity. The effects of such spontaneous excitatory activity on antigravity muscle tone can be tempered by activation of the inhibitory medullary reticular nuclei. Descending projections from the cerebral cortex to the brainstem represent a significant forebrain source of relative control over the two portions of the reticulospinal tract. This cortico-reticulospinal route emphasizes the point that some of the descending brainstem motor pathways provide an indirect way for more rostral levels of the motor system hierarchy to influence spinal lower motor neurons (see Figure 10-3). As noted in Chapter 8, γ motor neurons are usually activated along with α motor neurons so that muscle spindles maintain their sensitivity to stretch even when the muscle is shortened during contraction. This α-γ co-activation is a principle common to the excitation of lower motor neurons by upper motor neurons. Under certain circumstances, however, it appears that this process can be dissociated, such that the γ motor neuron–mediated sensitivity of the muscle spindle, and thus the sensitivity of the stretch reflex, can be adjusted apart from extrafusal muscle contraction. Although the reticulospinal tract participates in α-γ co-activation of lower motor neurons, it appears to be strongly associated with the ability to regulate γ motor neuron activity independently. It is likely that this ability of the reticulospinal tract to modulate independently the sensitivity of the stretch reflex underlies its significant role in adjusting antigravity muscle tone. The vestibulospinal tract originates from cell bodies in the vestibular nuclear complex, which lies primarily in the medulla, just ventral to the fourth ventricle (see Figure 11-8). This complex consists of several subnuclei that receive their principal synaptic input from the eighth cranial nerve fibers carrying sensory input from the vestibular apparatus of the inner ear (see Chapter 11). The vestibular apparatus provides sensory information about the position of the head with respect to gravity and about acceleration of the head through space, thus indicating body position and disturbances of balance. The vestibular nuclear complex also receives significant input from the cerebellum, but not from forebrain levels of the motor system hierarchy. As in the reticulospinal tract, axons of the vestibulospinal tract synapse within medial regions of the spinal cord gray matter that primarily control the axial and proximal extensor musculature (see Figure 10-3). Also like the reticulospinal tract, vestibulospinal tract axons collectively project to virtually all rostrocaudal levels of the spinal cord. When the vestibular apparatus detects a disturbance of balance, it excites the antigravity musculature in an attempt to counteract the disturbance. Although the vestibulospinal tract principally functions to produce compensatory adjustments to postural disturbances, it seems to make some contribution to antigravity muscle tone as well. The tectospinal tract is a medial brainstem motor pathway that is principally involved in reflex orientation of the head toward environmental stimuli. The cells of origin of the tectospinal tract are located in the superior colliculus of the midbrain (often called rostral colliculus in quadrupeds; see Figure 10-3). Like the other two medial brainstem motor pathways to the spinal cord, the tectospinal tract axons synapse within medial regions of the spinal cord gray matter that primarily control the axial and proximal musculature. However, these axons only project as far as the upper cervical regions of the cord. This is consistent because the tectospinal tract principally controls the musculature that moves the head. The superior colliculus processes visual, auditory, and somatosensory information about the relative position of stimuli in the environment with respect to the organism. The superior colliculus can also control rapid reflex movements (saccades) of the eyes to the stimulus. The tectospinal tract is involved in producing a movement of the head toward the stimulus that corresponds with the rapid eye movement so that the animal’s gaze is fixated directly on the stimulus.
The Central Control of Movement
The Central Nervous System Structures That Control Movement Have a Hierarchical Organization
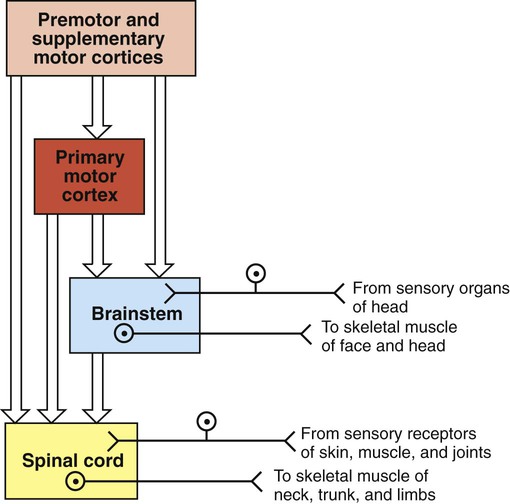
The Spinal Cord Is the Most Caudal and Simplest Level of the Movement Control Hierarchy
Brainstem Upper Motor Neuron Pathways Are the Source of All Descending Motor System Input to the Spinal Cord, Except for One Other Major Pathway
Medial and Lateral Descending Brainstem Motor Pathways Respectively Control Proximal Muscles of Posture and More Distal Muscles of Skilled Movement
The Reticulospinal and Vestibulospinal Tracts Are Medial Brainstem Motor Pathways Important for Keeping the Body Upright Against the Pull of Gravity
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


