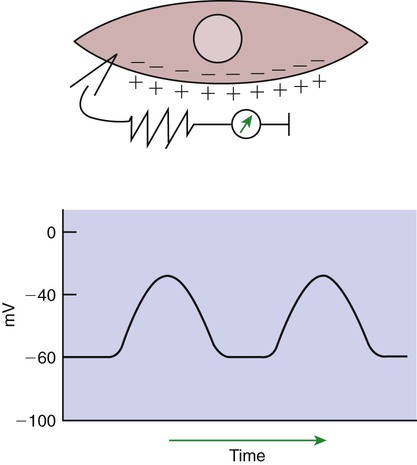
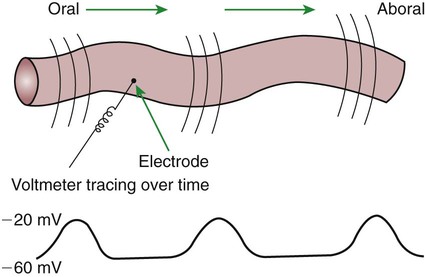
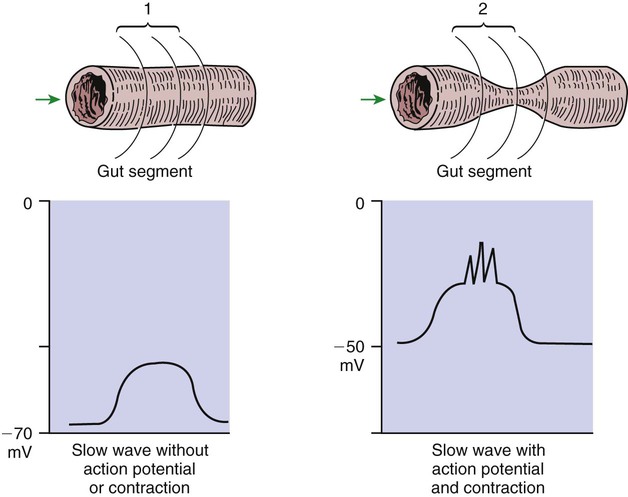
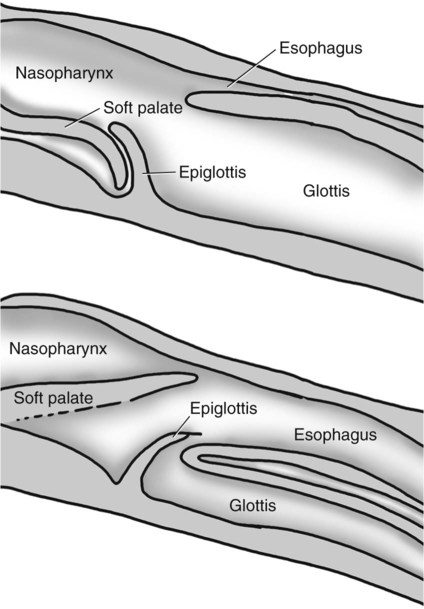
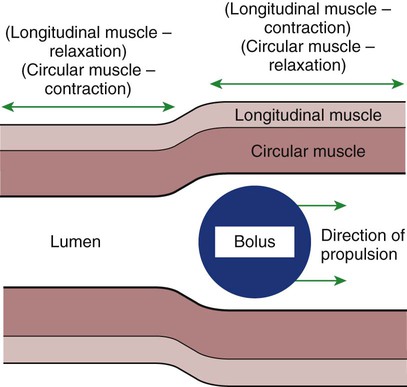
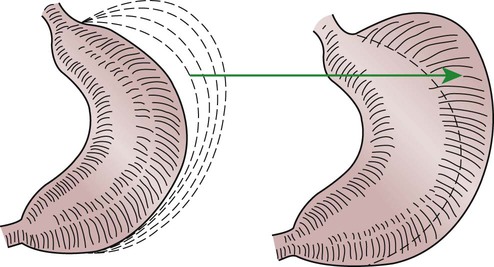
1. Slow waves of electrical depolarization are a unique feature of gut smooth muscle. 2. When slow waves reach sensitized smooth muscle cells, action potentials and contraction result. 3. Coordinated motility enables the lips, tongue, mouth, and pharynx to grasp food and propel it down the gastrointestinal tract. 4. Motility of the esophagus propels food from the pharynx to the stomach. 5. The function of the stomach is to process food into a fluid consistency and release it into the intestine at a controlled rate. 6. The proximal stomach stores food awaiting further gastric processing in the distal stomach. 7. The distal stomach grinds and sifts food entering the small intestine. 8. Control of gastric motility differs in the proximal and distal stomach. 9. The rate of gastric emptying must match the small intestine’s rate of digestion and absorption. 10. Between meals, the stomach is cleared of indigestible material. 11. Vomiting is a complex reflex coordinated from the brainstem. 12. Motility of the small intestine has digestive and interdigestive phases. 13. The ileocecal sphincter prevents movement of colon contents back into the ileum. 14. Motility of the colon causes mixing, retropulsion, and propulsion of ingesta. 15. The colon is an important site of storage and absorption in all animals. 16. Despite large anatomical differences in the colons of herbivores compared to omnivores and carnivores, there are similarities in motility. 17. The anal sphincter has two layers with separate innervation. 18. The rectosphincteric reflex is important in defecation. 19. Major differences between avian and mammalian digestive systems include, in birds, both the lack of teeth and the separation of gastric functions into distinct anatomical regions. The first level of control of GI motility lies in the intrinsic electrical properties of the smooth muscle mass. These electrical properties consist of spontaneously undulating waves of partial depolarization that sweep over the gut smooth muscle. The origin of this electrical activity is from specialized smooth muscle cells referred to as the interstitial cells of Cajal (ICC). The ICC form an interconnecting lattice of cells that surrounds the circular and longitudinal layers of muscle over the entire length of the gut. These cells are very similar in structure and function to the Purkinje cells of the heart. The ICC exhibit rhythmical and spontaneous oscillation in their transmembrane electrical potentials, as illustrated in Figure 28-1. They are connected to one another and to cells of the general smooth muscle mass by tight junctions or nexuses. These connections allow for the flow of ions from cell to cell. The resulting ionic movements lead to the propagation of waves of partial cell membrane depolarization across large numbers of cells. Within the ICC, fluctuations in intracellular calcium concentrations appear responsible for the spontaneous changes in membrane polarization. Figure 28-1 illustrates the concept of a fluctuating membrane potential in a single ICC. The property of spontaneous electrical rhythmicity, in combination with their electrical connection to the smooth muscle mass, imparts to the ICC their role as electrical “pacemakers” of the gut. In GI smooth muscle cells, the baseline membrane potential is usually –70 to –60 millivolts (mV). Under the influence of the ICC, the membrane potential fluctuates from this baseline level by as much as 20 to 30 mV. Thus, under resting conditions, the depolarization is only partial, and the membrane potential never reaches 0 mV. The smooth muscle cells are connected to the ICC and to each other by nexuses, allowing the changes in membrane potential to be spread, or propagated, over large areas of muscle. The ICC initiate these changes and thus determine their origin and direction of propagation. Under normal conditions in the small intestine, changes in membrane potential begin high in the duodenum and are propagated aborally (away from the mouth) along the length of the small intestine (Figure 28-2). These aborally moving waves of partial depolarization are called slow waves or the basic electrical rhythm of the gut. In the dog, slow waves occur about 20 times per minute in the small intestine. In the stomach and colon, slow waves occur less frequently, about five times per minute. However, the slow waves are present throughout the smooth muscle portions of the GI tract. The frequency of slow waves varies among domestic species, but their presence does not vary. Slow waves have an important relationship with muscle contractions, but they are not the direct stimuli for contractions. Slow waves are constantly passing over GI smooth muscle, whether it is actively contracting or not. GI smooth muscle cells, as with other muscle cells, contract in association with action, or spike, potentials. These potentials are characterized by complete depolarization of the membrane for a short time, in contrast to the slow waves, which are characterized by incomplete depolarization (see Chapter 4). Action potentials in the GI smooth muscle occur only in association with slow waves. Thus the presence of slow waves is necessary but not sufficient to cause muscle contractions. When slow waves pass over an area of smooth muscle without eliciting action potentials, no contractions occur. When slow waves pass over an area of smooth muscle and action potentials are superimposed on the slow waves, gut muscle contracts. Control and coordination of smooth muscle activity is achieved by influencing the likelihood that action potentials will be superimposed on slow waves. Such control is a function of peptides and regulatory substances produced by the ENS and enteric endocrine and paracrine cells. Smooth muscle control and coordination are achieved by modulation of the baseline electrical potential in the smooth muscle cells. Peptides and other regulatory molecules from ENS neurons or endocrine/paracrine cells are released in the vicinity of the smooth muscle cells, affecting membrane ion channels and influencing the baseline membrane potential (see Chapter 27 for a discussion of gut peptides and other regulatory molecules). Excitatory molecules elevate the baseline (bring it closer to zero), and inhibitory molecules lower the baseline (make it more negative). The position of the baseline influences how close the overall potential will come to 0 mV at the crest of a slow wave. When the membrane potential of a smooth muscle comes close to zero, action potentials occur and muscle contracts (Figure 28-3). Regulatory molecules (neurocrines, paracrines, and hormones) that are excitatory elicit smooth muscle contraction by elevating the baseline, whereas inhibitory substances inhibit muscle contraction by lowering the baseline. The involuntary actions of the swallow reflex occur primarily within the pharynx and esophagus. The pharynx is the common opening of both the respiratory and the digestive tract. The major physiological function of the pharynx is to ensure that air, and only air, enters the respiratory tract and that food and water, and only food and water, enter the digestive tract. The involuntary portion of the swallow reflex is the action that directs food into the digestive system and away from the upper airway. This reflex involves the following series of highly coordinated actions (Figure 28-4). Breathing stops momentarily. The soft palate is elevated, closing the pharyngeal opening of the nasopharynx and preventing food from entering the internal openings of the nostrils. The tongue is pressed against the hard palate, closing off the oral opening of the pharynx. The hyoid bone and larynx are pulled forward; this action pulls the glottis under the epiglottis, blocking the laryngeal opening. Concurrently, the arytenoid cartilages constrict, further closing the opening of the larynx and preventing the movement of food into the respiratory system. When all openings to the pharynx are closed, a wave of muscular constriction passes over the walls of the pharynx, pushing the bolus of food toward the opening of the esophagus. As the food reaches the esophagus, the upper esophageal sphincter relaxes to accept the material. In terms of motor activity, the esophagus may be viewed as consisting of an upper sphincter, body, and lower sphincter. The upper esophageal sphincter is called the cricopharyngeal muscle. This muscle and the upper end of the esophagus are attached to the cricoid cartilage of the larynx. When deglutition is not taking place, the muscle compresses the end of the esophagus against the cartilage of the larynx, tightly closing the upper esophageal opening. During deglutition the cricopharyngeal muscle relaxes and the larynx is pulled forward. The ventral portion of the upper end of the esophagus is attached to the larynx and the dorsal portion to the cervical spine. Because of these attachments, the forward motion of the larynx in conjunction with the relatively fixed nature of the cervical spine tends to pull open the upper esophageal orifice passively (see Figure 28-4). The body of the esophagus serves as a relatively simple conduit, rapidly transferring food from the pharynx to the stomach. Food is propelled through the esophagus by propulsive movements known as peristalsis. Peristalsis consists of a moving ring of constriction in the wall of a tubular organ. In the esophagus, these rings start at the cranial end and progress toward the stomach. The rings reduce or obliterate the esophageal lumen, thus pushing the bolus of food ahead of them in much the same manner as a person would push material out of a soft rubber tube by stripping it with the fingers. In addition to the constriction of the circular muscles, there may be some contraction of longitudinal muscles just ahead of, or aboral to, the ring of circular muscle contraction. This longitudinal muscle activity increases the size of the esophageal lumen to accommodate the advancing food bolus (Figure 28-5). Peristalsis is a universal type of GI propulsive motility that exists at all levels of the gut. Among animals there is tremendous diversity in the anatomy and motility patterns of the stomach. The following discussion applies best to the animals with the simplest stomachs, such as the dog and cat, but is probably also a reasonable description of the activity of the somewhat more complex stomachs of the pig, horse, and rat. The complex motility patterns of the ruminant stomach are discussed in Chapter 31. The major muscular activity in the proximal region of the stomach is of a weak, continuous-contraction nature. These tonic contractions tend to shape the gastric wall to its contents and provide gentle propulsion of material into the distal stomach. The major muscular reflex of the proximal stomach is adaptive relaxation (Figure 28-6). This reflex is characterized by relaxation of the muscles as food enters the stomach. Because of this relaxation, the stomach can dilate to accept large quantities of food without an increase in intraluminal pressure. Thus the proximal stomach serves as a food storage area. Because of the rather passive muscular activity of the proximal stomach, little mixing occurs there. In fact, food boluses tend to become layered in the stomach in the order in which they were swallowed. As the stomach empties, tension on the wall of the proximal stomach increases slightly, pushing food distally in the stomach, where it can be processed for transport into the duodenum.
Motility Patterns of the Gastrointestinal Tract
Slow Waves of Electrical Depolarization Are a Unique Feature of Gut Smooth Muscle
When Slow Waves Reach Sensitized Smooth Muscle Cells, Action Potentials and Contraction Result
Coordinated Motility Enables the Lips, Tongue, Mouth, and Pharynx to Grasp Food and Propel It Down the Gastrointestinal Tract
Motility of the Esophagus Propels Food from the Pharynx to the Stomach
The Function of the Stomach Is to Process Food into a Fluid Consistency and Release It into the Intestine at a Controlled Rate
The Proximal Stomach Stores Food Awaiting Further Gastric Processing in the Distal Stomach
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Motility Patterns of the Gastrointestinal Tract
Only gold members can continue reading. Log In or Register to continue
