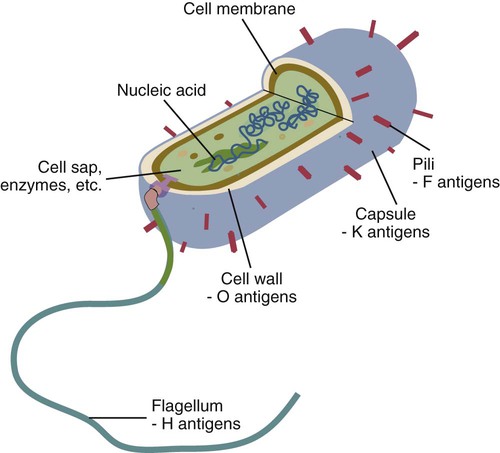
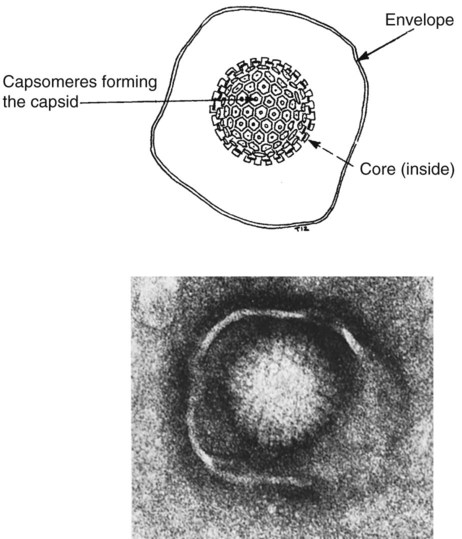
• The adaptive immune system is optimized to recognize microbial macromolecules. • The best antigens are therefore large, complex, stable, foreign proteins. • Small molecules of less than 5000 Da are usually poor antigens. • Small molecules may be made antigenic by linking them to large proteins. Small molecules used as antigens in this way are called haptens. • The cells of the adaptive immune system use receptors that can recognize specific areas on the surface of foreign molecules. These areas are called antigenic determinants or epitopes. Bacteria are single-celled prokaryotic organisms consisting of a cytoplasm containing the essential elements of cell structure surrounded by a lipid-rich cytoplasmic membrane (Figure 9-1). Outside the cytoplasmic membrane is a thick, carbohydrate-rich cell wall. The major components of the bacterial surface thus include the cell wall and its associated protein structures, the capsule, the pili, and the flagella. The cell wall of Gram-positive organisms is largely composed of peptidoglycan (chains of alternating N-acetyl glucosamine and N-acetyl muramic acid cross-linked by short peptide side chains) (see Figure 2-2). Gram-positive cell walls also contain lipoteichoic acids that are involved in the transport of ions across the cell wall. The cell wall in Gram-negative organisms, in contrast, consists of a thin layer of peptidoglycan covered by an outer membrane consisting of a lipopolysaccharide. Most of the antigenicity of Gram-negative bacteria is associated with the lipopolysaccharide. This consists of an oligosaccharide attached to a lipid (lipid A) and to a series of repeating trisaccharides. The structure of these trisaccharides determines the antigenicity of the organism. Many bacteria are classified according to this antigenic structure. For example, the genus Salmonella contains a major species, Salmonella enterica, that is classified into more than 2300 serovars based on antigenicity. These polysaccharide antigens are called O antigens. The outer cell wall lipopolysaccharides of Gram-negative bacteria bind to toll-like receptors (TLRs) and other pattern-recognition receptors and induce the production of inflammatory cytokines when an animal is infected. These cytokines cause a fever and sickness, so bacterial lipopolysaccharides are also called endotoxins. Viruses are very small organisms that can grow only inside living cells. They are thus “obligate,” intracellular parasites. Viruses usually have a relatively simple structure consisting of a nucleic acid core surrounded by a protein layer (Figure 9-2). This protein layer is termed the capsid, and it consists of multiple subunits called capsomeres. Capsid proteins are good antigens, highly capable of stimulating antibody formation. Some viruses may also be surrounded by an envelope containing lipoproteins and glycoproteins. A complete viral particle is called a virion. When a virus infects an animal, the virion proteins are processed and trigger adaptive immune responses. Viruses, however, are not always found free in the circulation but live within cells, where they are protected from the unwelcome attentions of antibodies. Indeed, viral nucleic acid can be integrated into a cell’s genome. In this situation, the viral genes code for new proteins, some of which are expressed on the surface of infected cells. These proteins, although they are synthesized inside an animal’s own cells, can still bind to antigen receptors and provoke adaptive immunity. These newly synthesized foreign proteins are called endogenous antigens to distinguish them from the foreign antigens that enter from the outside and are called exogenous antigens. Nucleated cells, such as leukocytes, possess hundreds of different protein molecules on their surface. These proteins are good antigens and readily provoke an immune response when injected experimentally into a different species. These surface molecules are classified by the CD system (Chapter 4). Other cell surface proteins may provoke an immune response (such as graft rejection) if transferred into a genetically different individual of the same species. The cell surface proteins that trigger graft rejection are called histocompatibility antigens. Histocompatibility antigens are of such importance in immunology that they warrant a complete chapter of their own (Chapter 11).
Antigens
Triggers of Adaptive Immunity
Microbial Antigens
Bacterial Antigens
Viral Antigens
Nonmicrobial Antigens
Cell Surface Antigens
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


