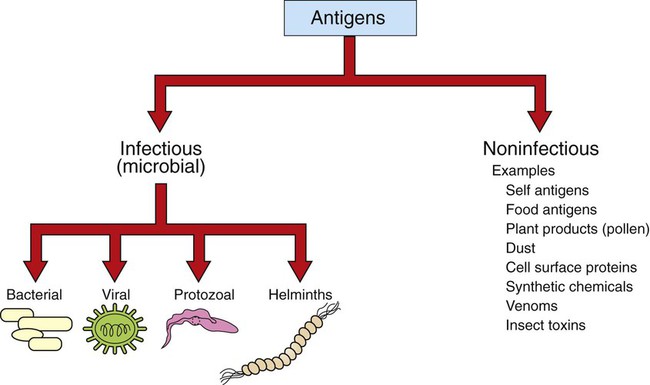
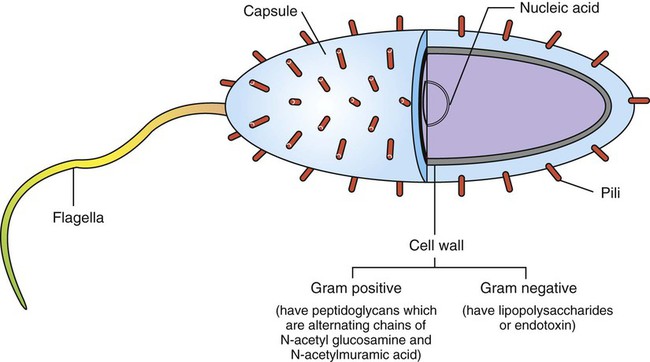
1. Antigens (or immunogens) stimulate immune cells to induce an immune response. 2. The degree of immune response depends on several characteristics of the antigen. Body’s Defense Against Invading Antigens 1. Both nonimmune and immune mechanisms defend against invading antigens. 2. A first line of defense includes physical and chemical barriers such as the skin and internal body fluids. 3. A second line of defense consists of phagocytic cells of the myeloid and macrophage-monocyte lineages. 4. Macrophage-derived cytokines can induce a variety of physiological processes to help combat infectious antigens. An antigen, or immunogen, is defined as any substance that is capable of stimulating immune cells (T and B cells) to induce an immune response. Antigens can be broadly divided into two large categories: (1) infectious (microbial) and (2) noninfectious (Figure 54-1). Infectious antigens include components that are derived from bacteria, viruses, protozoa, and helminths. Noninfectious antigens include those derived from “self” (autoantigens), food, plants, dust, or insect and animal venoms, as well as synthetic and cell surface proteins. Figure 54-2 shows the following antigenic structures of bacteria: • Bacterial cell wall. Cell walls of gram-positive bacteria differ from those of gram-negative bacteria. Gram-positive bacteria are composed of a thick layer of short chains of amino acids or peptides and carbohydrates (peptidoglycans). The cell wall of gram-negative bacteria has a thin layer of peptidoglycan and is largely composed of lipopolysaccharides, which are potent endotoxins. • Capsule. Certain bacteria produce a protective outer covering called a capsule, which is composed of polysaccharides. • Pili. These small, hairlike protein structures on some bacteria enable the bacteria to adhere to target host cells and transfer genetic information from one bacterium to another. • Flagella. Some bacteria possess flagella for mobility. Flagella contain a protein called flagellin, which can be antigenic. • Nucleic acids. Nucleic acids, such as bacterial deoxyribonucleic acid (DNA), tend to be antigenic because of differences in methylation compared with mammalian DNA. The antibodies against bacterial DNA tend to cross-react with the host’s DNA. • Foreign versus self antigens. Antigens that are considered to be foreign to the host tend to be highly antigenic. For example, if a horse is injected separately with antigens that are derived from a dog or from its own tissues, the horse will mount a strong immune response to the dog antigens, but not to its own (self) tissues. • Size. The size of an antigen also influences the level of immune response. Large antigens enable better processing by antigen-presenting cells (e.g., macrophages, dendritic cells) and subsequent presentation of antigenic peptides to lymphocytes for induction of an immune response. Examples of large antigens include bacterial and insect toxins, viral capsids, surface proteins on protozoa and helminths, and venoms. At the other extreme, very small antigens (e.g., small synthetic antigens, endogenous hormones, pesticides) tend to be ineffective in provoking an immune response. Very small antigens are inherently incapable of inducing immune responses; however, when bound to a larger protein, they can be potent antigens. Such small compounds are referred to as haptens. A good example of a hapten is a poison ivy–derived chemical, urishiol, which readily combines with many proteins (e.g., skin proteins) to induce a vigorous immune response. • Biochemical structure and complexity. In general, proteins tend to be more antigenic than lipids or carbohydrates. Large size alone is insufficient to provoke a good immune response. For example, many sugars and lipids, even though large in size, are ineffective in inducing an immune response because they consist of simple repeating units (e.g., repeating sugars in starch), which lack complexity. Complex carbohydrates and lipids, on the other hand, as found in many microbes, are strong immunogens. Carbohydrates and lipids, when combined with protein to form glycoproteins and lipoproteins, respectively, have increased complexity and thus are good antigens. • Stability and degradability. For immune cells to respond, stability of an antigen is an important feature. Flexible antigens, such as flagellin in a bacterium, are poor immunogens. However, when stabilized and rendered less flexible, as done in vaccine preparations, flagellin tends to be a potent immunogen. For an immune response to be initiated, the antigen ingested by phagocytic cells (e.g., macrophages) must be degraded and broken down into small peptides. Lymphocytes (T cells) will only respond to the peptides and not to large, native molecules. Antigens such as steel pins or plastic heart valves, even though large and complex, are inert and not degradable and thus are not good antigens. Initially, most antigens are effectively handled by nonspecific defense mechanisms, such as impervious and formidable physical barriers (e.g., skin and other body surfaces) and antimicrobial body fluids (e.g., lysozymes in tears, saliva, gastric juices). These are considered the first line of defense and are discussed next. Should the antigen survive this “body armor,” phagocytic cells (e.g., neutrophils, macrophages-monocytes) and natural killer (NK, NK-T) cells can effectively eliminate the invading antigens. These cells ingest and destroy a wide range of antigens and thus are non–antigen-specific. These cellular defenses constitute a second line of the body’s defense. The body’s initial defense (physical, chemical, and phagocytic antigen-presenting cells; natural killer cells) constitute the innate immune system. The antigen-presenting cells closely interact with specific T and B cells to induce a specific immune response. Thus, specific immune responses by the “adaptive” immune system tend to be the last line of the body’s defense (see Chapter 55). Collectively, both nonimmune and immune mechanisms effectively counter invading microbes.
Antigens and Innate Immunity
Antigens
Antigens (or Immunogens) Stimulate Immune Cells to Induce an Immune Response
The Degree of Immune Response Depends on Several Characteristics of the Antigen
Body’s Defense Against Invading Antigens
Both Nonimmune and Immune Mechanisms Defend Against Invading Antigens
A First Line of Defense Includes Physical and Chemical Barriers such as the Skin and Internal Body Fluids
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


