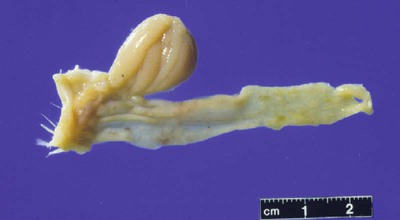
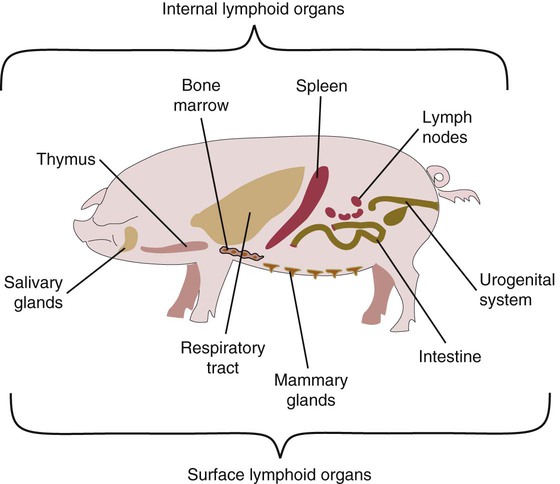
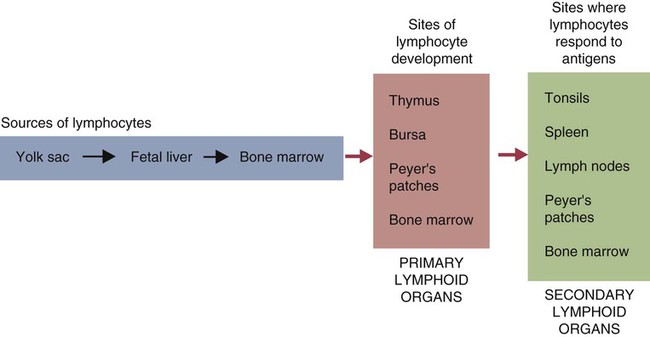
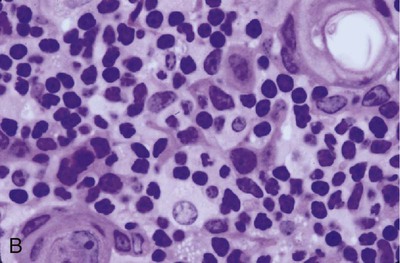
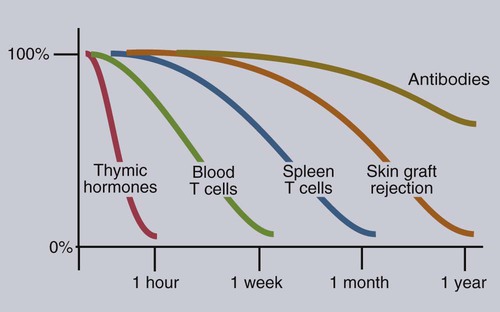
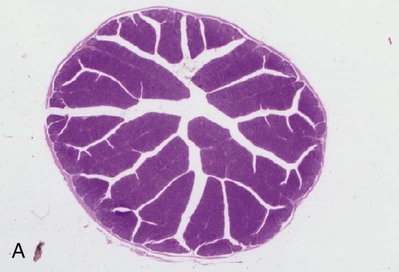
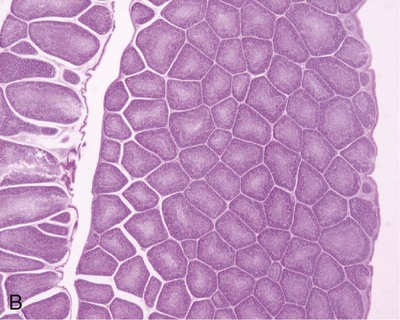
• Adaptive immunity is mediated by cells called lymphocytes that are found mainly within lymphoid organs. • Lymphocytes arise from stem cells in the bone marrow. • Lymphocytes mature within primary lymphoid organs. There are two types of lymphocyte: T cells and B cells. • T cells mature within the thymus. B cells mature within gastrointestinal lymphoid tissues, the bone marrow, or the bursa of Fabricius, depending on species. • If newly developed lymphocytes have receptors for self-antigens that could potentially cause tissue damage, they are killed before they can leave primary lymphoid organs. • Mature lymphocytes leave the primary lymphoid organs to reside in secondary lymphoid organs, where their role is to encounter and respond to foreign antigens. • The major secondary lymphoid organs include lymph nodes, spleen, bone marrow, and some Peyer’s patches within the intestine. Although antigens are captured and processed by dendritic cells, macrophages, and B cells, adaptive immune responses are actually mounted by cells called lymphocytes. Lymphocytes are the small round cells that predominate in organs such as the spleen, lymph nodes, and thymus (Figure 12-1). These are called lymphoid organs. Lymphocytes have antigen receptors on their surface and can therefore recognize and respond to foreign antigens. Lymphocytes are eventually responsible for the production of antibodies and for cell-mediated immune responses. The lymphoid organs must therefore provide an environment for efficient interaction among lymphocytes, antigen-presenting cells, and foreign antigens as well as sites where lymphocytes can respond optimally to processed antigens. Immune responses must be carefully regulated. Lymphocytes must be selected so that their receptors will only bind foreign antigens, and the response of each lymphocyte must be regulated so that it is sufficient but not excessive for the body’s requirements. The lymphoid organs may therefore be classified on the basis of their roles in generating lymphocytes, in regulating the production of lymphocytes, and in providing an environment for trapping foreign antigens, processing them, and maximizing the opportunity for processed antigens to encounter and interact with lymphocytes (Figure 12-2). The organs that regulate the development of lymphocytes are called primary lymphoid organs. Lymphocytes fall into two major populations called T cells and B cells, based on the primary organ in which they mature. Thus, all T cells mature in the thymus. B cells, in contrast, mature within different organs depending on species. These include the bursa of Fabricius in birds, the bone marrow in primates and rodents, and the intestinal lymphoid tissues in rabbits, ruminants, and pigs. The primary lymphoid organs all develop early in fetal life. As animals develop, newly produced, immature lymphocytes migrate from the bone marrow to the primary lymphoid organs, where they mature (Table 12-1). The primary lymphoid organs are not sites where lymphocytes encounter foreign antigens, and they do not enlarge in response to antigenic stimulation. Table 12-1 Comparison of Primary and Secondary Lymphoid Organs The thymus consists of lobules of loosely packed epithelial cells, each covered by a connective tissue capsule. The outer part of each lobule, the cortex, is densely infiltrated with lymphocytes (or thymocytes), but the inner medulla contains fewer lymphocytes, and the epithelial cells are clearly visible (Figure 12-3). Within the medulla are also found round, layered bodies called thymic or Hassall’s corpuscles. These contain keratin, and the remains of a small blood vessel may be found at their center. In cattle these corpuscles may contain immunoglobulin A (Chapter 16). An abnormally thick basement membrane and a continuous layer of epithelial cells surround the capillaries that supply the thymic cortex. This barrier prevents circulating foreign antigens from entering the cortex. No lymphatic vessels leave the thymus. As an animal ages, the thymus shrinks and is gradually replaced by fat. However, the aged thymus still contains small amounts of lymphoid tissue and remains functionally active. The functions of the thymus are best demonstrated by studying the effects of its removal in rodents. These effects depend on the age of the animal. For example, mice thymectomized within a day of birth become susceptible to infections and may fail to grow. These animals have very few circulating lymphocytes and cannot reject foreign organ grafts because they lose the ability to mount cell-mediated immune responses (Table 12-2). In contrast, adult thymectomy has no immediate obvious effect. But if these mice are monitored for several months, the number of blood lymphocytes and their ability to mount cell-mediated immune responses gradually decline. This suggests that the thymus remains functional in adults, but there is a reservoir of long-lived thymus-derived cells that must be exhausted before the effects of adult thymectomy become apparent (Figure 12-4). Table 12-2 Effects of Neonatal Thymectomy and Bursectomy Within the thymus, cell functions are regulated by a complex mixture of cytokines and small peptides collectively known as thymic hormones. These include peptides variously called thymosins, thymopoietins, thymic humoral factor, thymulin, and the thymostimulins. Thymulin is especially interesting because it is a zinc-containing peptide secreted by the thymic epithelial cells, and it can partially restore T cell function in thymectomized animals. Zinc is an essential mineral for the development of T cells. Consequently, zinc-deficient animals have defective cell-mediated immune responses (Chapter 38). Hassall’s corpuscles play a functional role in regulating thymic activity since they express a growth factor called thymic stromal lymphopoietin. This molecule activates thymic dendritic cells that can stimulate regulatory T cells and controls the positive selection process. The bursa of Fabricius is found only in birds. It is a round sac located just above the cloaca (Figure 12-5). Like the thymus, the bursa reaches its greatest size in the chick about 1 to 2 weeks after hatching and then shrinks as the bird ages. It is very difficult to identify in older birds. Like the thymus, the bursa consists of lymphocytes embedded in epithelial tissue. This epithelial tissue lines a hollow sac connected to the cloaca by a duct. Inside the sac, folds of epithelium extend into the lumen, and scattered through the folds are round masses of lymphocytes called lymphoid follicles (Figure 12-6). Each follicle is divided into a cortex and a medulla. The cortex contains lymphocytes, plasma cells, and macrophages. At the corticomedullary junction there is a basement membrane and capillary network on the inside of which are epithelial cells. These medullary epithelial cells are replaced by lymphoblasts and lymphocytes in the center of the follicle. Specialized neuroendocrine dendritic cells of unknown function surround each follicle.
Organs of the Immune System
Primary Lymphoid Organs
PRIMARY
SECONDARY
Origin
Ectoendodermal junction or endoderm
Mesoderm
Time of development
Early in embryonic life
Late in fetal life
Persistence
Involutes after puberty
Persists in adults
Effect of removal
Loss of lymphocytes
No or minor effects
Response to antigen
Unresponsive
Fully reactive
Examples
Thymus, bursa, some Peyer patches
Spleen, lymph nodes
Thymus
Structure
Function
FUNCTION
THYMECTOMY
BURSECTOMY
Numbers of circulating lymphocytes
Disappear
No effect
Presence of lymphocytes in T-dependent areas
Disappear
No effect
Graft rejection
Suppressed
No effect
Presence of lymphocytes in T-independent areas
Minor depletion
Disappear
Plasma cells in lymphoid tissues
Minor drop
Disappear
Serum immunoglobulins
Minor drop
Major drop
Antibody formation
Minor effects
Major drop
Thymic Hormones
Bursa of Fabricius
Structure
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Organs of the Immune System
Only gold members can continue reading. Log In or Register to continue
