Chapter 8 Vestibular system
Key Points
 Vestibular input helps maintain positions of the eyes, neck, trunk and limbs that are appropriate for the head position.
Vestibular input helps maintain positions of the eyes, neck, trunk and limbs that are appropriate for the head position.


 Vestibular nuclei receive additional proprioceptive input from proprioceptors in neck muscles.
Vestibular nuclei receive additional proprioceptive input from proprioceptors in neck muscles.

 The output from the vestibular nuclei is modified by efferent fibres from the cerebellum.
The output from the vestibular nuclei is modified by efferent fibres from the cerebellum.

Structural and functional anatomy
The vestibular system is divided into peripheral and central components (Fig. 8.1); this segregation has clinical implications. The peripheral vestibular system consists of the vestibular components of the inner ear (receptors and axons of CN VIII), while the central vestibular system comprises the vestibular nuclei and their output.
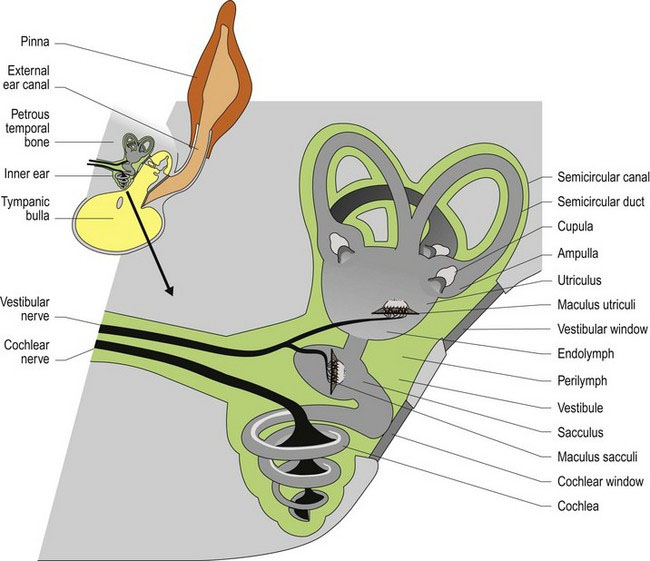
The inner ear is housed in the petrous temporal bone bilaterally. It contains a number of canals and cavities forming the bony labyrinth. On each side there are three semicircular canals, the bases of which connect to the bony vestibule. Ventral to the vestibule is the spiral cochlear canal that houses the auditory receptors (see Chapter 10). The semicircular canals arise from the vestibule, curve around and connect back to the vestibule. At one end of those connections the canal is dilated to form the ampulla. The canals and cavities are lined with membranes forming the membranous labyrinth. Perilymph is the fluid located between the membrane and the bone while endolymph fills the membranous labyrinth.
Static position, linear acceleration and deceleration of the head
Located within the vestibule are expanded portions of the labyrinth, called the utriculus and the sacculus (Fig. 8.1). These membrane-bound portions of the vestibular system each have a region called a macula; this is where the hair cells are located. The stereocilia and kinocilium of each cell project into an overlying gelatinous matrix called the otolithic membrane. Embedded in the membrane are calcareous crystals called otoliths that, having sufficient mass, move under the effect of gravity, pulling on the otolithic membrane and causing deflection of the stereocilia and kinocilium. The macula in the utriculus is oriented in the horizontal plane, whereas in the sacculus it is oriented vertically. Thus no matter what position the head is in, gravity will be acting on one or both sets of otoliths causing deflection of the cell processes on the hair cells and stimulating the sensory nerves at their base. The stimulation is constant due to the ongoing effect of gravity on the mass of the otoliths. Linear acceleration or deceleration will also affect otolith position (Fig. 8.2).

Angular acceleration and deceleration of the head
The three semicircular canals and their membranous ducts are located at right angles to each other (in the x, y and z directions) with the anterior and posterior canals being vertically oriented and the horizontal canal being laterally oriented. The semicircular ducts on each side of the head function as synergic pairs. Turning the head in one direction will cause endolymph to flow towards the ampulla on that side, but away from the ampulla in the paired duct on the opposite side. This causes stimulation of hair cells on the first side, but inhibition on the opposite side. Thus the input received by the vestibular nuclei of the brainstem is unequal. Unequal input from paired semicircular canals is interpreted as head movement. Note that if there is disease in the vestibular system on one side, then even at rest the brain will receive uneven input and thus, it is perceived that the animal’s head is moving. The uneven input from the two sides will be interpreted as head movement inducing the reflex changes in posture and eyeball position/movement (see Clinical dysfunction) that result in clinical signs of vestibular disease.
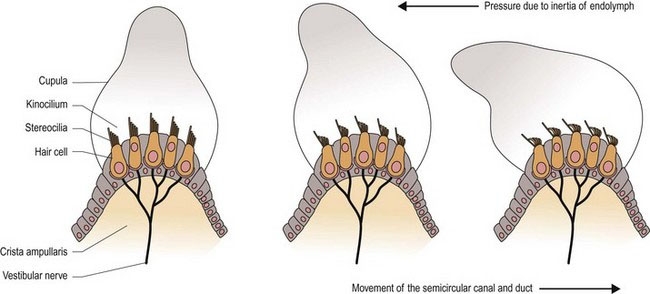
As the head moves, so will the petrous temporal bone, and the associated semicircular ducts and hair cells (Fig. 8.3 and 8.4). However, due to inertia, movement of the endolymph inside the ducts lags behind. The endolymph presses on the cupula, which deflects the stereocilia towards or away from the kinocilium, resulting in stimulation or inhibition of the vestibular neurons, respectively.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


