Body heat is produced by metabolism and is also gained from the environment. In homeotherms, internal temperature is normally maintained within a narrow range (37–40°C) by integrated neurophysiologic mechanisms that balance heat production and heat loss. Thermoregulation is the process by which the internal temperature is regulated to maintain body temperature within this thermoneutral zone. For example, during heat exposure or exercise, when heat gain occurs, the thermoregulatory system will provoke mechanisms for heat loss such that the rise in internal (or core) body temperature is mitigated. It is the most important regulation system in homeothermic animals.1,2 Core body temperature is a dynamic equilibrium between factors that add or remove heat. This balance is maintained by integration of mechanisms that vary the body’s rate of heat production, alter the transfer of heat to the periphery (e.g. skin), and regulate evaporative cooling. The hypothalamus contains the central coordinating center for the various processes of thermoregulation. Specialized heat- and cold-sensitive neurons within the preoptic and anterior nuclei of the hypothalamus act as a thermostat to initiate thermoregulatory adjustments to deviations from normal body temperature. Input from thermal receptors in the skin (as well as the abdominal viscera, large veins, and spinal cord) provide information to the hypothalamus. This input, compared and integrated with detected changes in the temperature of blood perfusing the anterior hypothalamus, forms the basis for the initiation of thermoregulatory adjustments.3,4 The greatest disequilibrium in heat balance occurs during exercise. Conversion of chemical energy (i.e. stored substrates) to mechanical energy (e.g. muscular contraction) is inefficient, with approximately 70–80% of the total chemical energy released as heat rather than physical work.4–6 As such, the rate of metabolic heat production increases markedly with the onset of exercise and is accompanied by increases in muscle and core body temperatures. The increase in core temperature provokes activation of heat dissipatory mechanisms such that increases in body temperature are mitigated. However, whether balance between heat gain and heat loss can be re-established will depend on the duration and intensity of exercise and the efficiency of heat dissipation. The latter is primarily influenced by ambient conditions but also modified by hydration state and by physiological adaptations (e.g. conditioning, heat acclimatization) in heat dissipatory mechanisms.4,7,8 The body has a basal level of energy expenditure or basal metabolic rate (BMR) that is lowest when environmental temperature range is within a thermoneutral zone. This BMR minimizes energy expenditure required to maintain normal body temperature. The BMR is altered by numerous factors, both internal and within the environment, which increase or decrease energy expenditure. Metabolic rate is normally lowered during sleep and increases with any form of work or stress. Perturbations of BMR such as exercise, fever, catecholamine release, feed consumption (thermic effect of feed), or dealing with a cold environment can therefore affect heat balance. In mammals, there have been extensive investigations of the relationship between bodyweight and heat production,5,9 which have demonstrated that resting heat production is proportional to body mass to the power of 0.75 in adults but not in growing animals. There are, however, differences between species and it could be expected that within a species such as the horse with extensive variation in size and weight based on breed, lighter breeds may have a significantly lower resting heat production when compared to heavier breeds.10 Heat production in horses is influenced by dietary factors, including the quantity and quality of feed and water intake. Replacing hay in the diet with grains has been demonstrated to decrease heat production,11 with a further reduction in heat production possible with fat supplementation.12 When exposed to hot or cold environments, water restriction and dehydration can also reduce heat production. During exercise, the workload or speed is the main determinant of the rate of heat production. Other factors such as the weight of rider and tack, and the nature of the terrain and footing will also contribute to the overall workload.13,14 Heat production during exercise can be estimated from oxygen consumption data: where In contrast to most other large domestic species in which skeletal muscle comprises 30–40% of total bodyweight, half the total bodyweight of the Thoroughbred is working muscle. This higher percentage of bodyweight from muscle contributes to the horse’s higher mass-specific Radiative heat transfer occurs when waves of electromagnetic radiation are emitted or absorbed at the skin surface. As body temperature is normally higher than the environment, there is a net loss of radiative heat energy at the skin surface. However, in hot ambient conditions, when the temperature of objects in the environment exceeds skin temperature, radiant heat energy is absorbed from the surroundings. Under these conditions, the only avenue for heat loss is evaporative cooling. A gain of radiant heat energy also occurs via direct (or reflected) sunlight. It has been suggested that solar radiation can contribute up to 15% of the heat gain in horses during exercise in sunny conditions.15 For the horse, the most important mechanism for heat loss is evaporative cooling including the evaporation of sweat from skin surfaces and water from the respiratory tract. The efficacy of this mechanism is dependent upon the extent of the vapor pressure gradient between body surface and environment. Calculations of the estimated heat loss are based on the latent heat of vaporization of water (from a liquid to a vapor – 598 kcal [2501 kJ] for each gram of water at 0°C).16 After accounting for possible variations in the thermodynamic properties of sweat when compared to water, it is estimated that the evaporation of 1 L of sweat from the skin surface will dissipate approximately 580 cal (2428 kJ or 2.4 MJ) of body heat.17 The quantity of heat (~2.4 MJ) dissipated in association with the evaporation of 1 L of sweat in thermoneutral conditions is approximately equivalent to the heat generated by 2 min of high-intensity exercise or 6 min of moderate-intensity exercise. In optimum conditions (i.e. when relative humidity is low) the evaporation of sweat is a very efficient mechanism of heat loss and can account for as much as 65% of total heat loss during exercise. However, several environmental factors will influence the efficacy of evaporative heat loss. These include ambient temperature and relative humidity, the extent of the vapor pressure gradient between the skin surface, and the rate of air movement.18–20 At high ambient humidity, the vapor pressure gradient between the body surface and the environment narrows, thereby constraining evaporative cooling and increasing the rate of heat storage. The extensive surface area of the respiratory tract also provides a mechanism for heat dissipation. This process relies upon the difference in vapor pressure between the inspired air and that of the epithelial surface of the respiratory tract. The external nares of the horse contribute considerable surface area for heat exchange. Similarly, the extensive surface area of the upper respiratory tract, including the internal nares and nasal turbinates provide an environment in which air entering the nasal passages contacts the highly vascularized epithelium of the upper respiratory tract. Horses, like humans, lack a carotid rete, a vascular heat exchanger that can serve to cool blood shortly before it enters the brain. However, horses have a unique anatomical arrangement by which their internal carotid arteries are enveloped by a pair of air-filled guttural pouches. Studies by Baptiste et al21 suggest exercising horses utilize these guttural pouches to cool blood en route to the brain. It is surmised that this heat loss from the upper respiratory tract contributes to selective brain cooling, whereby the temperature of blood reaching the brain is lower when compared with that measured in mixed venous (pulmonary artery) blood or within skeletal muscle.22 The extent of selective brain cooling in the horse is questionable as not all studies to date have reported a lower hypothalamic vs carotid artery temperature during and following exercise.23 In only a limited number of species, including some bovidae, primates and equidae, is the sweat gland primarily a thermoregulatory organ.11,24 In the horse, sweat glands are present in both haired and relatively hairless skin with regional variation in the density of glands that is not dependent on the presence of a haircoat. Structurally, the gland is similar to that of many other domestic species, consisting of a fundus and a duct connecting the fundus with the skin surface. Throughout most of its length, the duct lining is composed of two layers, with a single layer of keratinocytes lining the duct at the skin surface. Located within the dermis, the fundus is lined by an inner layer of secretory epithelium interspersed with myoepithelial cells and surrounded by a fenestrated sheath of fibrocytes that encloses a layer of connective tissue.25 Although sweating appears to be under sympathetic nervous control, there is no evidence of direct sympathetic innervation of the sweat gland. Rather, sweating appears to occur via humoral stimulation of β2-adrenergic receptors on sweat glands. As a result, sweating can be initiated by epinephrine (adrenaline) release in advance of any stimulus related to an increase in core temperature. However, Scott et al26 reported peak sweating rates in standing horses in response to infusion of epinephrine (adrenaline) were significantly lower than rates previously measured by these investigators during exercise, which supported the concept of a dual control of sweating that integrates hormonal and neural input. As most studies to date have investigated sweating rate and composition in Thoroughbred horses, the degree to which sweating rate and composition varies between breeds is unknown. To date, it appears that the basic composition of sweat is similar between breeds. Spooner et al27 noted that sweat sodium concentration measured in Arabian horses performing endurance exercise was lower than values for sweat sodium measured in previous studies but this have been a reflection of the low sodium diet imposed for this study. Equine sweat, unlike that of humans, is isotonic to slightly hypertonic relative to plasma.18,22,28 Equine sweat is also unique in having a high protein content, a consequence of the presence of latherin. This surface-active, non-glycosylated protein significantly reduces water surface tension, aiding the translocation of sweat fluid through the haired coat to facilitate rapid evaporation.29 Sweat ion concentrations are largely a reflection of sweating rate and therefore are subject to alteration based on environmental conditions and exercise intensity (Table 41.1). Although there is some variation in sweat ion composition of equine sweat, individual differences in sweat composition do not appear to be as extensive in horses when compared to human athletes. Epinephrine (adrenaline) infusion will produce a more dilute sweat and may account for the less concentrated sweat produced during high-intensity exercise when compared to low-intensity exercise.18,22,30 Table 41.1 Sweating rate, ion concentrations, and osmolality at two exercise intensities and three different ambient conditions after 10 minutes of exercise CD = cool, dry (room temperature [T] = 20°C, relative humidity [RH] = 45–55%); HD = hot, dry (T = 32–34°C, RH = 45–55%); HH = hot, humid (T = 32–34°C, RH = 80–85%); High = exercise at 90% of maximum oxygen consumption ( Data from McCutcheon LJ, Geor RJ, Hare MJ, et al. Sweat composition: comparison of collection of methods and effects of exercise intensity. Equine Vet J Suppl 1995;18:279–84. McCutcheon LJ, Geor RJ, Hare MJ, et al. Sweating rate and sweat composition during exercise and recovery in ambient heat and humidity. Equine Vet J Suppl 1995;20:153–7. (Refs 18, 132). During exercise, an increase in body temperature as a result of metabolic heat production is the primary stimulus for sweating. Normally, sweating is initiated at a specific core temperature and continues in proportion to the increase in core temperature.31 demonstrated that increases in sweating rate at three different exercise intensities (40%, 65%, and 90% of Skin blood flow is substantially increased by the opening of capillary beds that are normally bypassed by arteriovenous anastamoses that connect arteries directly to veins. The increase in blood flow through the vascular beds of the skin allows heat to be lost to the environment via convection and direct radiation of heat from the skin surface. The efficacy of transfer of heat by convection and radiation varies according to the rate of air movement across the skin (wind speed) and the gradient of skin temperature to environmental temperature. Increased skin blood flow also provides the latent heat for vaporization of sweat and as well as the fluid required for sweat production. The attempt to maximize blood flow for increased activity and thermoregulation is also reflected in decreased splanchnic and adipose tissue blood flow.32 Greater oxygen demand increases respiratory rate and respiratory blood flow and both activities will enhance the extent of evaporative cooling by the respiratory system. Sweating rates of ~20 to 55 mL/m2/min have been measured on the necks and backs of horses exercising on a treadmill in a laboratory.31,33–35 Assuming a body surface area of 4.5 to 5.0 m2 for a 500 kg horse, these sweating rates correspond to fluid losses of 6–15 L per hour. This estimate of hourly sweat fluid loss is in agreement with sweat rates calculated on the basis of the decrease in body mass during prolonged exercise under field conditions.36 When expressed in terms of sweating rate per unit area of skin, these sweating rates are two- to three-fold greater than those reported for human subjects. At any given point in time during exercise, core body temperature reflects the balance between heat production and dissipation. Soon after the onset of exercise, the rate of heat production greatly exceeds the rate of heat dissipation such that there is a rapid increase in muscle temperature.37 During short-term, high-intensity exercise (e.g. racing), the rate of heat production will exceed the rate of heat loss throughout exercise and body temperature will continue to increase until the cessation of exercise. In this circumstance, a large proportion of the metabolic heat load will be dissipated during the recovery period. Conversely, during more prolonged low- to moderate-intensity exercise in temperate ambient conditions, activation of heat dissipatory mechanisms progressively attenuates the rate of rise of body temperature. Eventually, the rate of heat loss increases sufficiently to balance metabolic heat production, allowing a near steady-state core temperature to be achieved.37 Not surprisingly, the thermal response to exercise is affected by the ambient conditions. As environmental temperature increases, the thermal gradient between the skin and the environment is reduced, and sensible heat loss (i.e. convective and radiative heat transfer) is impaired. When ambient temperature exceeds skin temperature (>35–36°C), the gradient for heat transfer is reversed and the body gains heat from the environment. If humidity is low, a decrease in sensible heat loss can be offset by an increase in sweating rate and evaporative cooling. As humidity rises, the gradient between skin and ambient dew point is reduced and evaporative heat loss is also impaired. The decrease in sweat evaporation is manifested by excessive wetting of the skin surface and drippage of sweat from the body. Sweat that drips from the body only removes 5% to 10% of the heat that can be dissipated by evaporation of sweat. Therefore, during exercise under conditions of high ambient heat and humidity, the rate of heat dissipation may be inadequate to prevent the progressive rise in body temperature. The impact of the environment on the rate of rise of core body temperature in exercising horses is depicted in Figure 41.2. The rate of heat storage when exercising in hot, humid conditions may be more than twice the rate occurring during exercise at the same intensity in cool, dry conditions.20,38–40 Increased demands for respiratory heat loss are reflected by an increase in respiratory rate and during and after exercise. Kohn and Hinchcliff41 reported a 20% to 25% increase in the respiratory rate of horses during speed and endurance tests in hot when compared to cool conditions. In laboratory experiments, an approximately two-fold increase in post-exercise respiratory rate has been observed in horses under hot, humid when compared to cool, dry environmental conditions.20,38 When ponies were exposed to heat (41°C dry bulb temperature), there were three-fold increases in respiratory rate and blood flow to tissues of the upper respiratory tract,42 reflecting the role of the respiratory system in heat dissipation. Similarly, during moderate-intensity (~30% An important consequence of the impairment to heat dissipation during exercise in the heat is a decrease in the time to attainment of a critical upper limit in core body temperature. In humans, it is clearly established that time to exhaustion in trained subjects during exercise in the heat is inversely related to the initial level of body temperature and directly related to the rate of heat storage.43 That the lowering of core temperature prior to the start of exercise44 or cooling the body during the period of exercise45 will delay attainment of the critically high body temperature and extend exercise duration is further evidence for the relationship between body temperature and exercise performance. Several factors may contribute to a decrease in performance when exercise is undertaken in hot versus cool conditions. These include the effects of hyperthermia on brain and muscle function, and compromise of cardiovascular and respiratory function. There appears to be a critical body temperature above which mammals will not continue to exercise voluntarily, likely a protective mechanism to protect the human or animal from reaching tissue temperatures that harm cell function.40 Thus, a more rapid attainment in critical body temperature will translate to a reduction in exercise duration.46 In trained humans exercising over a range of work intensities in the heat, voluntary fatigue occurs at a core (esophageal) temperature of 39.7–40.0°C.43,47–49 Elevations in core body temperature (either through passive heat exposure or exercise) also increase serotonergic (5-hydroxytryptamine; 5-HT) and dopaminergic pathway activity. There is evidence that both pathways contribute to the vasodilatory component of thermoregulatory control and to the release of neuroendocrine hormones that can contribute to the behavioral (motivational) response to exercise. Intrinsic differences in these pathways have been proposed as one mechanism contributing to differences in heat tolerance between individuals. For example, differences in receptor density or neurotransmitter release to a given stimulus could produce variations in the thermoregulatory or behavioral responses to a given thermal load.50 Furthermore, following heat acclimation procedures that enhance thermoregulatory mechanisms and reduce the rate of heat storage during exercise, the core temperature at the onset of fatigue is unchanged.48,51 Measurements of central blood (pulmonary artery) temperature in horses during heavy exercise have demonstrated that fatigue occurs as blood temperature approaches 42.5–43°C;31,52 muscle temperature may reach 44–45°C during such high-intensity exercise. Hypothalamic blood temperature, on the other hand, is approximately 1°C lower than central blood temperature in horses during heavy exercise in moderate ambient conditions.22 The difference between the temperatures in these two regions provides evidence for the existence of a mechanism for selective brain cooling in the horse. Particularly during exercise in the heat, the onset of fatigue at some critical upper limit in brain temperature may represent a mechanism to avoid heat stroke. Human studies, dating back to the work of Asmussen and Boje,53 have indicated that a moderately elevated, but steady-state core body temperature is advantageous to muscle function and to the dissociation of oxygen from red blood cells within muscle tissue. The increase in muscle temperature acts on glycolytic and glycogenolytic enzymes, altering flux rate through these pathways.54,55 This Q10 effect is accentuated during exercise in the heat. However, at high muscle temperature (>46°C) deleterious structural and functional alterations in skeletal muscle proteins can be induced.56 These proteins play essential roles in mitochondrial respiration, regulation of calcium by the sarcoplasmic reticulum and the subsequent interactions of myosin and actin, and control of electrolyte movement across the sarcolemma.43,57 As a consequence, substantial detrimental alterations to skeletal muscle metabolism may occur with elevation of muscle temperature to this critical range. During exercise heat stress, circulatory adjustments must be regulated to maintain adequate blood flow to contracting muscle and to the thermoregulatory tissues, particularly the skin and the upper respiratory tract.17,58 Given a finite cardiac output, the increased demands for blood flow to these thermoregulatory tissues may compromise blood flow to skeletal muscle, thereby limiting oxygen delivery and, possibly, exercise duration. Studies in ponies have provided data on the effects of environmental heat load (41°C dry bulb) on the redistribution of cardiac output during exercise.31 Blood flow to the fore- and hindlimbs during moderate- (~30% A reduction in maximal aerobic power is another potential reason for decreased physical performance during exercise in the heat. A reduction in peak oxygen uptake has been demonstrated in human subjects exercising in the heat.59–61 When non-heat acclimatized horses performed an incremental treadmill exercise test in hot, humid conditions (temperature 30°C, relative humidity 75%), peak expired minute ventilation, oxygen uptake and oxygen pulse were significantly lower and plasma lactate concentrations higher when compared to exercise in temperate conditions (15°C, 55% relative humidity).62 Although the mechanism of the reduction in peak oxygen uptake in horses exercising in heat and humidity has not been determined, it is possible that alterations in breathing strategy that favor heat loss from the upper respiratory tract (dead space ventilation) and compromise of skeletal muscle blood flow result in decreased oxygen uptake and delivery. Regardless of mechanism, such a reduction in peak oxygen uptake is likely to impair performance relative to exercise performed in temperate conditions. The term ‘exercise heat tolerance’ refers to an ability to withstand high internal and external heat loads during exercise. In humans it is well recognized that high aerobic fitness and a period of acclimatization in the heat improves both physiological and psychological responses to the challenge of exercise in the heat.63 Adaptations in heat dissipatory mechanisms improve cardiovascular stability, decrease the rate of heat storage, and increase the duration of exercise before volitional fatigue. However, highly trained athletes are also able to tolerate higher levels of hyperthermia when compared to untrained individuals: that is, volitional fatigue occurs at a higher core temperature.64 Conversely, regardless of training or heat acclimatization, dehydration decreases exercise heat tolerance, as evidenced by an increase in the rate of heat storage and the development of volitional fatigue at a lower level of hyperthermia. There is now evidence that these physiological factors also modify the thermoregulatory capacity of horses. Specifically, conditioning and heat acclimatization improve tolerance to exercise in the heat, whereas dehydration adversely affects heat dissipation in horses during exercise. Other factors that can influence thermoregulation in exercising horses include coat color and the density of the hair coat. Coat color will affect the quantity of solar heat absorbed, while a long hair coat will limit evaporative heat loss. Breed-associated physiological traits have also been implicated in thermal tolerance and the extent of seasonal adaptation to environmental temperature.10,65–67 Physical training in a cool environment is broadly accepted to improve exercise heat tolerance. The extent to which heat dissipatory effector mechanisms are stimulated and the duration of that stimulus will determine the effectiveness of training in improving exercise-heat tolerance. Detectable changes in heat tolerance are evident in human athletes after 1–2 weeks of training but are much more substantial if regular training is sustained for 8–12 weeks.68–70 In human subjects, the stimulus must be sufficient to elevate core temperature by approximately 1.5 to 2.0°C for a minimum of 30 min/day.68,70,71 At a given percent of maximal oxygen uptake (
Thermoregulation and exercise-associated heat illnesses
Introduction
Heat production and dissipation
Heat production
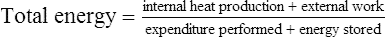
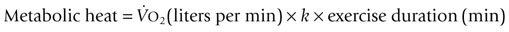
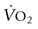 = oxygen consumption and k = amount of heat liberated per liter of oxygen consumed. Values for k range from 4.7 kcal to 5.1 kcal depending on the substrate oxidized (lowest value for pure fat oxidation, highest value for pure carbohydrate oxidation).9
= oxygen consumption and k = amount of heat liberated per liter of oxygen consumed. Values for k range from 4.7 kcal to 5.1 kcal depending on the substrate oxidized (lowest value for pure fat oxidation, highest value for pure carbohydrate oxidation).9
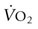 when compared to other athletic species, including man. Furthermore, a running horse uses a greater proportion of its body mass for locomotion than does a human performing running or leg cycling exercise. Thus, the mass-specific heat load for the exercising horse is as much as two- to three-fold higher compared to that of exercising humans. Despite a substantially higher rate of heat production in the horse, the ratio of surface area to body mass is approximately 50% less than that of humans (man = 1 : 35–40 m2/kg; horse = 1 : 90–100 m2/kg).13,14 As a result, the horse has a significantly smaller surface area over which to dissipate a relatively larger metabolic heat load and, at any given workload, must dissipate approximately four times more heat per unit of body surface area during exercise than human athletes. The disadvantage posed by a smaller surface area to body mass ratio can be partially offset by higher rates of cutaneous and respiratory heat loss. However, it is apparent that exercise is a considerable thermoregulatory challenge to the horse, with prolonged exercise representing one of the most demanding situations.
when compared to other athletic species, including man. Furthermore, a running horse uses a greater proportion of its body mass for locomotion than does a human performing running or leg cycling exercise. Thus, the mass-specific heat load for the exercising horse is as much as two- to three-fold higher compared to that of exercising humans. Despite a substantially higher rate of heat production in the horse, the ratio of surface area to body mass is approximately 50% less than that of humans (man = 1 : 35–40 m2/kg; horse = 1 : 90–100 m2/kg).13,14 As a result, the horse has a significantly smaller surface area over which to dissipate a relatively larger metabolic heat load and, at any given workload, must dissipate approximately four times more heat per unit of body surface area during exercise than human athletes. The disadvantage posed by a smaller surface area to body mass ratio can be partially offset by higher rates of cutaneous and respiratory heat loss. However, it is apparent that exercise is a considerable thermoregulatory challenge to the horse, with prolonged exercise representing one of the most demanding situations.
Mechanisms of heat transfer
Mechanisms of sweat formation
VARIABLE
CD HIGH
CD LOW
HD LOW
HH LOW
Sodium (mmol/L)
124.0 ± 6.7
116.7 ± 6.1
133.6 ± 2.3
130.6 ± 1.7
Potassium (mmol/L)
25.8 ± 2.1
32.6 ± 1.4
41.5 ± 1.1
28.1 ± 0.9
Chloride (mmol/L)
142.0 ± 5.6
144.3 ± 4.3
155.8 ± 3.2
149.5 ± 2.9
Osmolality (mOsm/kg)
313 ± 18
303 ± 6
339 ± 6
327 ± 5
Sweating rate (mL/m2/min)
40.4 ± 3.7
21.1 ± 5.2
32.8 ± 5.1
27.0 ± 6.2
 ); low = exercise at 50% of
); low = exercise at 50% of  .
.
 ) were closely related to elevations in carotid artery blood temperature (Fig. 41.1). Rate of rise in body temperature and the concentration of circulating catecholamines associated with different exercise intensities could also contribute to the determination of sweating rate.
) were closely related to elevations in carotid artery blood temperature (Fig. 41.1). Rate of rise in body temperature and the concentration of circulating catecholamines associated with different exercise intensities could also contribute to the determination of sweating rate.
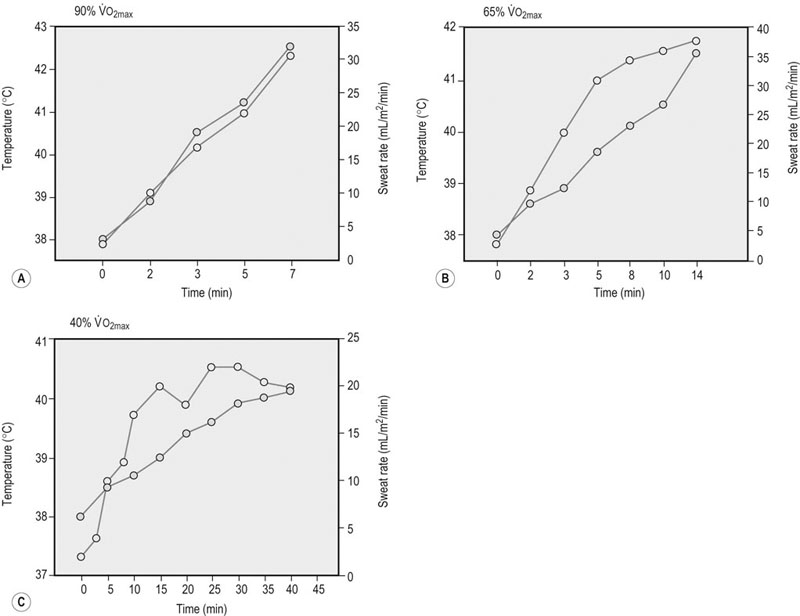
 ). Ambient temperature was 21–23.5°C. Adapted from Hodgson DR, McCutcheon LJ, Byrd SK, et al. Dissipation of metabolic heat in the horse during exercise. J Appl Physiol 1993;74(3):1161–70, with permission from the American Physiological Society.31
). Ambient temperature was 21–23.5°C. Adapted from Hodgson DR, McCutcheon LJ, Byrd SK, et al. Dissipation of metabolic heat in the horse during exercise. J Appl Physiol 1993;74(3):1161–70, with permission from the American Physiological Society.31
Thermoregulation during exercise
Effects of environmental heat load on exercise responses
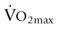 ) exercise respiratory rate was five-fold greater in hot than in thermoneutral conditions.32
) exercise respiratory rate was five-fold greater in hot than in thermoneutral conditions.32
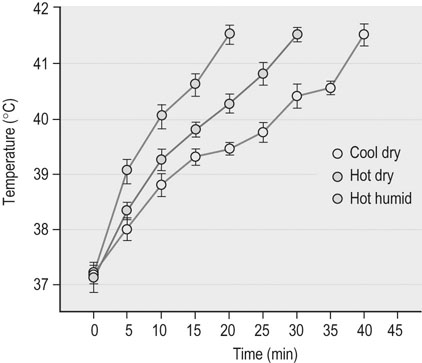
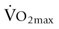 ) and high-intensity (~65%
) and high-intensity (~65% 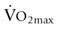 ) exercise was reduced by as much as 25–30% when compared to similar exercise in thermoneutral conditions. This reduction in muscle blood flow during exercise heat stress is likely to restrict performance and contribute to an early onset of fatigue.
) exercise was reduced by as much as 25–30% when compared to similar exercise in thermoneutral conditions. This reduction in muscle blood flow during exercise heat stress is likely to restrict performance and contribute to an early onset of fatigue.
Physiologic factors affecting thermoregulatory capacity
Conditioning
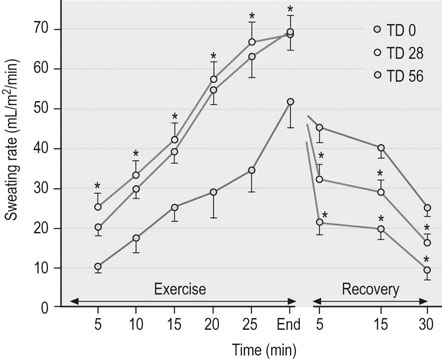
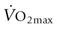 ), trained athletes have a higher metabolic rate and, hence, greater heat production at any given relative exercise intensity. Despite this higher metabolic rate, the trained individual is able to maintain a similar core temperature when compared to the untrained subject, indicating an enhanced ability to dissipate heat. Improved cutaneous blood flow and whole-body sweating have been measured in trained individuals and are the main contributions to more effective heat dissipation in human subjects.72 However, highly trained human athletes have been shown to have less extensive sweat fluid losses during exercise73,74 and there is evidence for similar sweating economy in horses following training.75 Specifically, eights weeks of moderate intensity treadmill conditioning resulted in a 1.6-fold increase in sweating sensitivity and an approximately 0.7°C decrease in sweating threshold in horses during exercise in hot, dry conditions (temperature 32–34°C, relative humidity 45–55%). Despite higher sweating rates for a given core temperature during exercise, decreases in recovery sweating rates resulted in an overall reduction in sweat fluid losses (Fig. 41.3).75
), trained athletes have a higher metabolic rate and, hence, greater heat production at any given relative exercise intensity. Despite this higher metabolic rate, the trained individual is able to maintain a similar core temperature when compared to the untrained subject, indicating an enhanced ability to dissipate heat. Improved cutaneous blood flow and whole-body sweating have been measured in trained individuals and are the main contributions to more effective heat dissipation in human subjects.72 However, highly trained human athletes have been shown to have less extensive sweat fluid losses during exercise73,74 and there is evidence for similar sweating economy in horses following training.75 Specifically, eights weeks of moderate intensity treadmill conditioning resulted in a 1.6-fold increase in sweating sensitivity and an approximately 0.7°C decrease in sweating threshold in horses during exercise in hot, dry conditions (temperature 32–34°C, relative humidity 45–55%). Despite higher sweating rates for a given core temperature during exercise, decreases in recovery sweating rates resulted in an overall reduction in sweat fluid losses (Fig. 41.3).75
Thermoregulation and exercise-associated heat illnesses
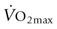 ) when compared to the resting state. For a 500 kg horse with a
) when compared to the resting state. For a 500 kg horse with a 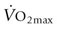 of 80 L/min, this equates to metabolic heat production in excess of 400 kcal/min (~1.3 MJ/min) of exercise. Production of this quantity of heat without any ability for heat dissipation would result in an increase in body temperature of approximately 1°C per minute during exercise. Although the rate of metabolic heat production is lower during endurance exercise, the overall heat load is substantially higher because of the longer work duration. For example, it has been estimated that the metabolic heat production of an endurance horse running at 8 m/s is about 150–200 kcal/min; if no heat was dissipated, this heat load would result in an increase in core temperature of approximately 21°C per hour. These hypothetical measurements emphasize that effective heat-loss mechanisms are crucial and serve to further underline the additional impact severe ambient conditions will impose on the horse’s ability to lose heat to the surrounding environment.
of 80 L/min, this equates to metabolic heat production in excess of 400 kcal/min (~1.3 MJ/min) of exercise. Production of this quantity of heat without any ability for heat dissipation would result in an increase in body temperature of approximately 1°C per minute during exercise. Although the rate of metabolic heat production is lower during endurance exercise, the overall heat load is substantially higher because of the longer work duration. For example, it has been estimated that the metabolic heat production of an endurance horse running at 8 m/s is about 150–200 kcal/min; if no heat was dissipated, this heat load would result in an increase in core temperature of approximately 21°C per hour. These hypothetical measurements emphasize that effective heat-loss mechanisms are crucial and serve to further underline the additional impact severe ambient conditions will impose on the horse’s ability to lose heat to the surrounding environment.