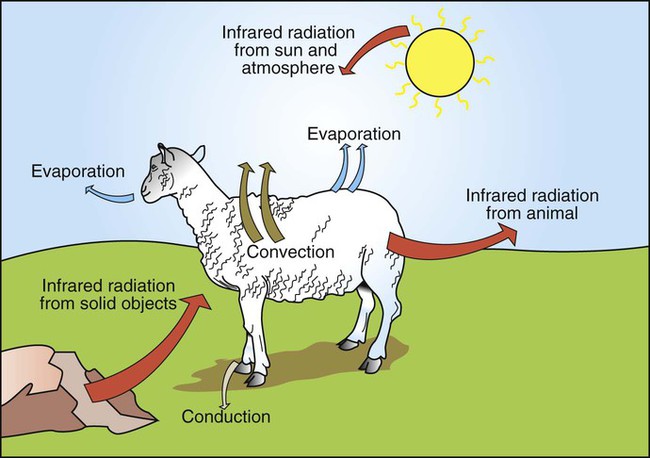
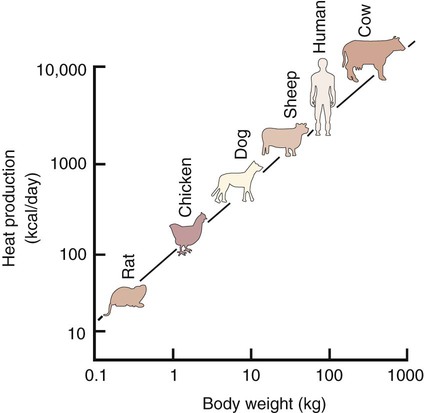
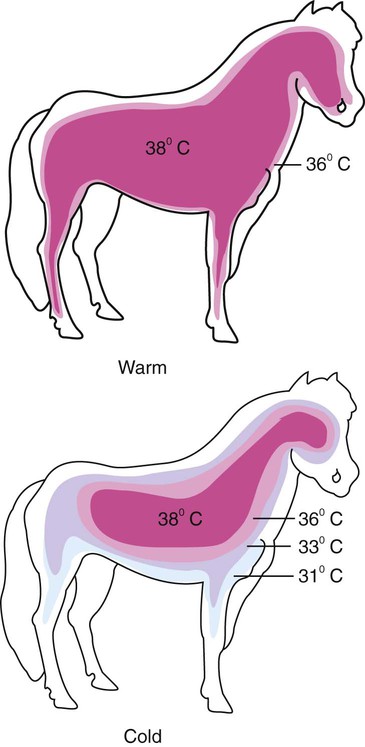
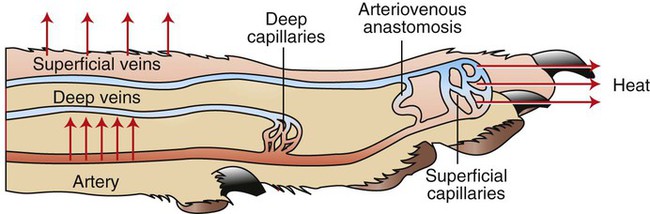
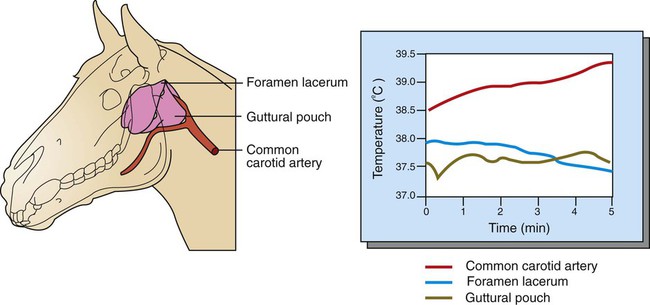
1. Temperature is a major factor affecting tissue function. 2. Homeotherms and poikilotherms use different strategies to regulate body temperature. 3. Body temperature depends on the balance between heat input and heat output. 1. Heat is a byproduct of all metabolic processes. 2. Shivering produces heat by muscle contraction. 3. Nonshivering thermogenesis is an increase in basal metabolic rate, caused especially by the oxidation of brown fat, to produce heat. 1. Because tissues are poor conductors, heat is most effectively transferred in the blood. 2. Countercurrent heat exchange mechanisms are used both to conserve and to lose heat. Heat Exchange with the Environment 1. Heat loss by convection occurs when the body warms air or water. 2. Heat loss by conduction occurs when the body is in contact with a cooler surface. 3. Heat loss by radiation occurs when infrared radiation emitted by the body is absorbed by cooler objects. 4. Heat loss by evaporation occurs when the water in sweat, saliva, and respiratory secretions is converted into water vapor. 1. Mammals and birds regulate the input and output of heat to maintain body temperature within a narrow limit. 2. Temperature-sensitive receptors are located in the central nervous system, the skin, and some internal organs. 3. Information from central and peripheral heat-sensitive neurons is integrated in the hypothalamus to regulate heat-losing or heat-conserving mechanisms. 1. The responses to heat stress are peripheral vasodilation and increased evaporative cooling. 2. The responses to cold stress are peripheral vasoconstriction, piloerection, and increased metabolic heat production by shivering and nonshivering thermogenesis. 3. Fever is an elevation of body temperature that results from an increase in the thermoregulatory set point. Heat Stroke, Hypothermia, and Frostbite 1. Heat stroke occurs when heat production or input exceeds heat output, so body temperature rises to dangerous levels. 2. Hypothermia occurs when heat output exceeds heat production, so body temperature decreases to dangerous levels. 3. Frostbite occurs when ice crystals form in the tissues of the extremities. Heat inputs to the body are from metabolism and from external sources (Figure 53-1). When food energy is ingested, heat is produced at all stages of the metabolic process. Eventually, all food energy is converted into heat, which is dissipated into the environment and radiated into space. Heat production by the body is related to metabolic rate. A basal metabolic rate is necessary to maintain the function of cells. During exercise, metabolic heat production can increase more than tenfold. If this heat is not dissipated to the environment, body temperature can increase to dangerous levels. Furthermore, this increase in body temperature increases metabolic rate, which further increases heat production. Table 53-1 shows the amount of heat produced by the metabolism of carbohydrates, fats, and proteins. The basal metabolic rate (BMR) is the rate of energy metabolism measured under minimal stress while the animal is fasting. BMR is greater in homeotherms than in poikilotherms because homeotherms need to generate heat to maintain body temperature. The BMR per kilogram of body weight is greater in smaller than in larger mammals (Figure 53-2). This is necessitated partly by the greater surface/volume ratio of smaller animals. The relatively greater surface area per kilogram body weight of small animals provides a larger area for heat loss. TABLE 53-1 Amount of Heat Produced by Metabolism of Major Food Types When animals are chronically exposed to cold, they develop the ability to increase metabolic heat production without shivering (nonshivering thermogenesis). This increase in metabolism is mediated through an increase in secretion of thyroid hormones and the calorigenic effects of catecholamines on lipids. Table 53-1 shows that fat metabolism is an effective way to produce heat. Brown fat is a specialized vascular, mitochondria-rich fat that is used to generate heat. It is widely distributed within the body of all mammals but is especially prevalent in small mammals where, combined with white fat it forms both subcutaneous and trunk depots. Cold stress–induced neural release of norepinephrine from the primarily adrenergic nerve supply activates β3-adrenergic receptors on adipocytes which increases fat metabolism to produce heat that is distributed around the body through the bloodstream. Furthermore, under cold stress white adipocytes can be converted to the brown adipocytes necessary for heat generation. Under conditions of heat stress, circulatory transfer of heat to the skin can be increased dramatically by two mechanisms. First, the arterioles of skin vascular beds dilate, which results in increased capillary blood flow. Second, arteriovenous anastomoses open in the limbs, ears, and muzzle. These two actions greatly increase the total blood flow to the periphery, and the increased heat delivery increases the temperature of the skin, which facilitates heat loss. Conversely, under cold stress, skin vascular beds vasoconstrict, and arteriovenous anastomoses close; thus skin and limb temperatures decrease. This results in reduced heat loss from the skin and in a gradient of temperatures along the limb (Figure 53-3). Under severe cold stress, the skin temperature of the extremities can approach ambient temperature. Interestingly, the lipids in the limb extremities have a lower melting point than those in the core, so fats do not solidify in extreme cold stress. When the environmental temperature is high, the blood perfusing the skin vascular beds returns to the body core through superficial veins from which heat is lost to the skin and air. Under cold conditions, limb blood flow returns to the core through deep veins that accompany arteries (Figure 53-4). Heat is transferred by countercurrent exchange from the warm arterial blood to the cooler venous blood and thereby returned to the core of the body. Some mammals, including humans and horses, do not possess a carotid rete and must rely on other thermoregulatory mechanisms to cool their brains during exercise. In the horse the guttural pouches may serve as such a mechanism. The guttural pouches contain air that is cooler than the arterial blood carried in the internal carotid artery. Because anatomically these guttural pouches surround the internal carotid arteries, heat is transferred from the blood to the air in the guttural pouches, thus protecting the brain from hyperthermia (Figure 53-5). In addition, the intracranial cavernous venous sinuses may assist in cooling the horse’s brain during exercise. This mechanism is thought to function in the same manner as the carotid rete, but less efficiently.
Thermoregulation
Body Temperature Depends on the Balance Between Heat Input and Heat Output
Heat Production
Heat Is a Byproduct of All Metabolic Processes
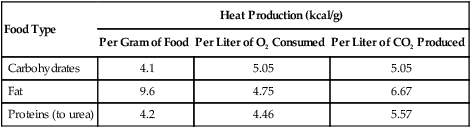
Food Type
Heat Production (kcal/g)
Per Gram of Food
Per Liter of O2 Consumed
Per Liter of CO2 Produced
Carbohydrates
4.1
5.05
5.05
Fat
9.6
4.75
6.67
Proteins (to urea)
4.2
4.46
5.57
Nonshivering Thermogenesis Is an Increase in Basal Metabolic Rate, Caused Especially by the Oxidation of Brown Fat, to Produce Heat
Heat Transfer in the Body
Because Tissues Are Poor Conductors, Heat Is Most Effectively Transferred in the Blood
Countercurrent Heat Exchange Mechanisms Are Used Both to Conserve and to Lose Heat
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Thermoregulation
Only gold members can continue reading. Log In or Register to continue
