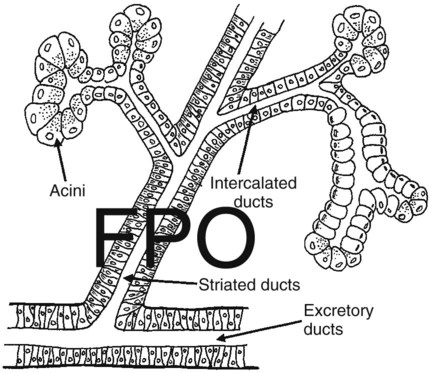
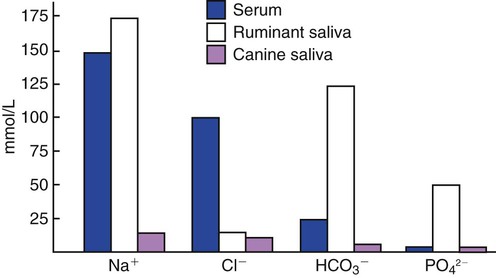
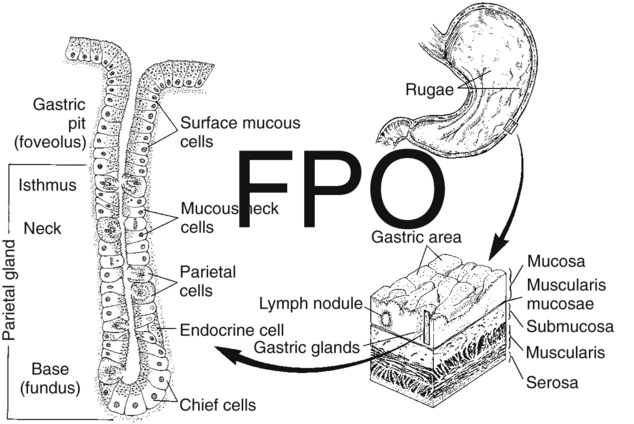
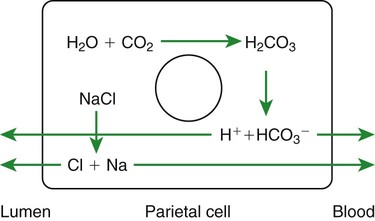
1. Saliva moistens, lubricates, and partially digests food. 2. Salivary secretions originate in the gland acini and are modified in the collecting ducts. 3. Salivary glands are regulated by the parasympathetic nervous system. 4. Ruminant saliva is a bicarbonate-phosphate buffer secreted in large quantities. 1. Depending on the species, there may be two general types of gastric mucosa: glandular and nonglandular. 2. The gastric mucosa contains many different cell types. 3. The gastric glands secrete hydrochloric acid. 4. Pepsin is secreted by gastric chief cells in an inactive form and is subsequently activated in the stomach lumen. 5. The parietal cells are stimulated to secrete by the action of acetylcholine, gastrin, and histamine. 1. Pancreatic exocrine secretions are indispensable for the digestion of the complex nutrients: proteins, starches, and triglycerides. 2. Acinar cells secrete enzymes, whereas centroacinar cells and duct cells secrete an electrolyte solution rich in sodium bicarbonate. 3. Pancreatic cells have cell surface receptors stimulated by acetylcholine, cholecystokinin, and secretin. 1. The liver is an acinar gland with small acinar lumina known as canaliculi. 2. Bile contains phospholipids and cholesterol maintained in aqueous solution by the detergent action of bile acids. 3. The gallbladder stores and concentrates bile during the periods between feeding. 4. Bile secretion is initiated by the presence of food in the duodenum and stimulated by the return of bile acids to the liver. The evaporative cooling function of saliva is covered in Chapter 53. The salivary gland is a typical acinar gland composed of an arborizing system of collecting ducts that end in cellular evaginations known as acini (Figure 29-1). The cellular epithelium of the acini is functionally distinct from that of the collecting ducts. Saliva is initially secreted into the lumen of the acini. The glandular cells lining the acini secrete water, electrolytes, enzymes, and mucus. As the newly formed saliva progresses through the collecting ducts, its composition is modified. The duct epithelium reabsorbs electrolytes, especially sodium and chloride, in a manner similar to that in the proximal tubules of the kidneys. The final product, saliva, is hypotonic and has a sodium concentration substantially less than that of extracellular fluid. The extent to which the acinar secretion is modified in the collecting ducts depends on the rate of saliva production. At high rates of salivary flow, there is little modification, which results in higher tonicity and electrolyte concentration, in comparison to low rates of flow. The normal composition of ruminant parotid saliva is quite different from the saliva of monogastric animals. Bovine and canine saliva are compared in Figure 29-2. Ruminant saliva is isotonic and, compared with blood serum, has high concentrations of bicarbonate and phosphate and a high pH. This well-buffered solution is necessary for neutralizing acids formed by fermentation in the rumen, and ruminants secrete it in enormous quantities. An adult cow may secrete 100 to 200 L of saliva per day. This volume is approximately equivalent to the extracellular fluid volume of most adult cattle. It is obvious that much of the water and electrolytes secreted in saliva must be reabsorbed rapidly and recirculated through the total body water, or the cow would die of dehydration. In abnormal circumstances, such as blockage of the esophagus, in which the flow of saliva is diverted from the gastrointestinal (GI) tract, cattle quickly become dehydrated and acidotic. Most domestic, monogastric animals have only glandular mucosa in the stomach, but horses and rats have an area in the proximal portion of their stomachs that is covered by nonglandular, stratified squamous epithelium. This area is visibly different from the glandular area, to which it adjoins with a sharp line of demarcation. The function of the nonglandular area of gastric mucosa is unclear. The nonglandular area may serve as a place where a small amount of fermentative (rumenlike) digestion could occur. Because there is little mixing activity in the proximal stomach, food in the nonglandular area would be protected from the secretions of the gastric glands. These acid secretions kill bacteria, and thus their presence would prevent fermentation. Fermentative digestion is discussed in detail in Chapter 31. The glandular mucosa of the stomach has frequent invaginations, or pores, known as gastric pits. The size of the pits is such that the pores leading into them can be seen with a hand-held magnifying glass. At the base of each pit is a narrowing, or isthmus, that continues into the opening of one or more gastric glands (Figure 29-3). Hydrogen ions for secretion come from the dissociation of intracellular carbonic acid (H2CO3), leaving a bicarbonate ion (HCO3–) in the cell for each H+ secreted into the lumen (Figure 29-4). Carbonic acid originates from water and carbon dioxide through the action of carbonic anhydrase, an enzyme found in high concentration in the gastric mucosa.
Secretions of the Gastrointestinal Tract
The Salivary Glands
Saliva Moistens, Lubricates, and Partially Digests Food
Salivary Secretions Originate in the Gland Acini and Are Modified in the Collecting Ducts
Ruminant Saliva Is a Bicarbonate-Phosphate Buffer Secreted in Large Quantities
Gastric Secretion
Depending on the Species, There May Be Two General Types of Gastric Mucosa: Glandular and Nonglandular
The Gastric Mucosa Contains Many Different Cell Types
The Gastric Glands Secrete Hydrochloric Acid
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


