Fig. 1.
Mean total daily number of M&M’s® earned and wasted each candy session for the five animals who wasted M&Ms® (left panel ) and the three animals who ate nearly all of the M&M’s® earned (right panel ) as a function of week of access to candy. Although small numbers of M&M’s® were wasted during the first 3.5 weeks of M&M’s® access, the number wasted were not accurately counted until week 4. Although shown sequentially, candy was available on Mondays, Wednesdays, and Fridays. Error bars on the data points represent ±1 SEM.
When Skittles® replaced M&M’s®, the baboons that were in the Wasters group (Fig. 2, left panel) and the Eaters group for M&M’s® (right panel) nearly doubled Skittles® deliveries between the first and third or fourth day of candy access, but then Skittles® deliveries remained stable for the remaining 8 weeks of candy access (15) for both groups. There was no significant difference between the M&M’s® Wasters and Eaters in the number of Skittles® earned or wasted. All baboons wasted few Skittles®. No baboon wasted food pellets.
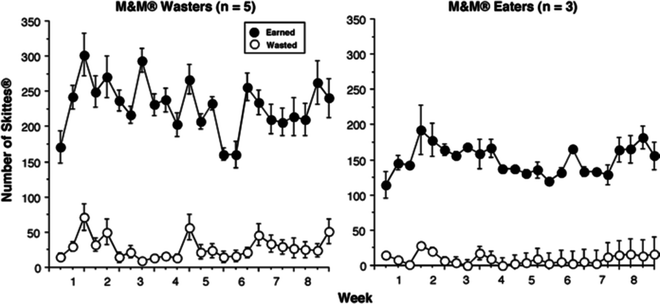
Fig. 2.
Mean total daily number of Skittles® earned and wasted each candy session for the five animals who wasted M&M’s® (left panel ) and the three animals who ate nearly all of the M&M’s® earned (right panel ) as a function of week of access to candy. Although shown sequentially, candy was available on Mondays, Wednesdays, and Fridays. Error bars on the data points represent ±1 SEM.
In summary, when given periodic opportunities to “forage” for a highly-palatable food item, free-feeding nonhuman primates worked for and consumed a large quantity of the preferred item. When candy was available, the number of candies consumed in the first meal of the day was similar to or larger than the total number of food pellets consumed over the rest of the day. Due to the greater caloric content of candy, baboons also consumed more calories on days that candy was available. Candy intake increased during the first week of candy access, but then remained stable across the period of access to candy.
Candy meals, based on a priori definition, lasted longer than pellet meals at the same time of day. Individuals with binge-eating disorder increase their caloric intake generally by having a longer meal (16, 17). All eight baboons ate one less food pellet meal on the days candy was available, thereby reducing total food pellet intake on the days when candy was available. Clinical data indicate that when humans eat a large binge meal, that meal substitutes for a small meal rather than being an additional meal that day (18–20). These similarities between human and nonhuman primates suggest that the present procedures may provide a model for large meals.
Five (three males and two females) of the eight baboons developed the unexpected behavior of earning then not fully eating M&M’s®, i.e., they did not eat the chocolate center. This wasting pattern generally occurred toward the end of the M&M’s® meal. Minimal tasting and wasting behavior occurred when baboons had access to Skittles®. This may have been due to the fact that Skittles® have five flavors while M&M’s® have only one flavor (six colors). Perhaps the wasting behavior, which only occurred in the laboratory when sugar-coated M&M’s® were available, reflects normal behavior observed when baboons are in a dense patch of preferred food. Of course, Skittles® are also coated in sugar, but they have a fruit-flavored, not chocolate-flavored interior.
The large candy meals in the current procedure parallel the consumption of excessive amounts of food within a single meal that is a defining characteristic of disordered eating in patients with bulimia nervosa and binge-eating disorder.
2.3 Effects of Pharmacological Manipulations on Binge Eating
The current procedure provides a behavioral baseline for evaluating potential pharmacological interventions (e.g., fluoxetine (21)) for these disorders. The specificity of an intervention on eating of a large meal of preferred items compared to “normal” eating can be evaluated by comparing the effects of drugs on days when a preferred food is available, and eaten during a large meal, to days when only food pellets are available, and eaten in multiple smaller meals. Drug effects are determined by administering a dose of drug prior to the first meal of the day, which in our case was available when the computers were restarted at 0900 hours.
We initially looked at a serotonergic and a dopaminergic drug based on evidence that these two neurotransmitters affect different aspects of feeding behavior. An increase in serotonin, as caused by dexfenfluramine (22), has been hypothesized to be vital for the development of satiation (see (23) for a review). An increase in dopamine, as caused by amphetamine (24), has been hypothesized to decrease hunger. Four doses of each drug plus placebo were tested, with each dose–response function requiring approximately 3 weeks for determination: placebo was tested on 25% of the test days to control for conditioned responses. At each dose–response determination, half of the baboons were tested with the smaller doses first and half were tested with the larger doses first. Dose order was systematically varied within and across baboons. Doses were given intramuscularly at 0900 hours when the computers were restarted for the day. At that time, completion of the first 30 min seeking interval resulted in the initiation of the first meal of the day that was either a Skittles® candy meal or a food pellet meal. Latency to the first candy or food delivery provided a measure of food seeking and was defined as the number of minutes between 0900 hours and the first food or candy delivery; the minimal interval was 30 min, including the duration of the seeking interval. Separate dose–response functions were determined when candy was available as the first meal of the day followed by food pellets meals and when food pellet meals were only available.
Dexfenfluramine did not affect the latency to the candy meal, but produced dose-dependent decreases in candy intake during the candy meal (Fig. 3; (25)). By contrast, dexfenfluramine dose-dependently increased the latency to the food pellet meal and decreased the number pellets consumed in the first food pellet meal. Amphetamine dose-dependently increased the latency to the food pellet and candy meals and decreased the number pellets and candies consumed in the first meal (26).
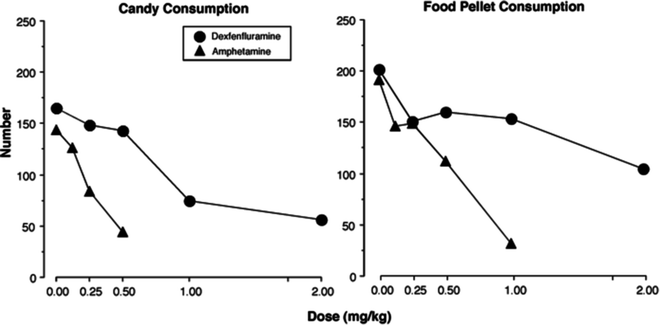
Fig. 3.
Mean number of Skittles® consumed during the candy meal (left panel ) and mean total number of food pellets (right panel ) consumed over the entire session as a function of dose of d-amphetamine and dexfenfluramine (N = 8: four female and four male baboons).
The absence of a significant effect of dexfenfluramine on candy seeking, coupled with a decrease in candy taking, supports the concept that dexfenfluramine, presumably by increasing serotonin, increases satiety. The data obtained with food pellets are a bit more difficult to align with the satiation hypothesis. Perhaps dexfenfluramine increased the satiety value of the food already consumed, thereby increasing the latency to the first food pellet meal. This effect was not seen on candy days, as animals were more motivated to respond to candy than to food. This assumes baboons learned the sequence of candy on Mondays, Wednesdays, and Fridays. Regardless, this dissociation between candy and food pellet seeking and taking was not observed for amphetamine, arguing that different behavioral mechanisms were involved for the two drugs. Thus, the behavioral baseline of 3 days access to a single candy meal before food pellet meals and 4 days access to only food pellet meals allowed for the investigation of the behavioral specificity of pharmacological agents.
The number of rewards earned during the seeking phase of the candy meal and food pellet meals was not affected by the drugs, indicating that responding during the 30-min intervals was not a sensitive measure of commodity seeking. This suggests that a simpler approach to providing food or candy meals could be used to provide a behavioral baseline. For example, the second taking component could be used alone.
3 Notes
One may be tempted to simplify the approach by providing monkeys free-access, i.e., a generous amount of the food in the home cage, to a supply of preferred food on an intermittent basis and determine intake by weighing the food before and after a limited period of access. When given free access to food, monkeys are wasteful in that they will pick and eat little bits of food and throw other bits away, not unlike the tasting and wasting described above. By adding a response cost, even a minimal one, e.g., ten lever pulls that take 2–3 S to accomplish, monkeys are generally less wasteful. The addition of the operant requirement allows for better measurement of intake (see (27) for an excellent history of the development of operant techniques to study feeding behavior). Although not presented, we collected data on rate of lever pulling and have examined the pattern of cumulative intake within a meal.
< div class='tao-gold-member'>
Only gold members can continue reading. Log In or Register to continue
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


