Fig. 28.1
Serial paraffin sections prepared by in vivo cryotechnique and stained with hematoxylin-eosin (HE, a) or immunostained for albumin (Alb, b), immunoglobulin G 1 (IgG1 , c), or IgM (d) show that in late primary follicle in living mouse ovaries, multiple layers of follicular cells surround the oocyte. Inside the blood vessel s , the clear immunoreactivity of the three serum protein s is observed (a–d, white arrowheads). Immunoreactivity of albumin (b, black arrowhead) and IgG1 (c, black arrowhead) is clear in the theca layers and interstitium of ovary, but immunostaining for IgG1 is weaker inside ovarian follicles (c, black arrow). Immunostaining for IgM is decreased in the interstitium and theca layers (d, black arrows) compared with that in blood vessels (d, white arrowheads) and hardly detected in the follicles (d, white arrow). Bars, 50 μm (The figure was modified from Zhou et al. [14])
28.3 Characteristics of BFB for Permselectivity
Serial ovarian sections with follicles in different stages were prepared with IVCT and immunostained to compare distributions of the three serum protein s including albumin (69 kDa), IgG1 (150 kDa), and IgM (900 kDa) (Fig. 28.1) [14]. In a late primary follicle with a stratified layer of granulosa cells around the oocyte, blood vessel s contained immunoreactivities of all three proteins. While IgG1 immunoreactivity was decreased inside the primary follicle, immunoreactivities of albumin and IgG1 were also observed in the interstitium (Fig. 28.1). To the contrary, strong immunostaining for IgM was observed inside blood vessels but reduced in theca layers, interstitium, and follicles. Therefore, large molecules such as IgG1 and IgM appear to be blocked around the follicles or the blood vessels, but small albumin easily enters ovarian follicles.
In primordial follicles composed of a primary oocyte and a single layer of flat follicular cells surrounding the oocyte, blood vessel s around those follicles immunostained for the three serum protein s (Fig. 28.2) [14]. The albumin immunostaining was positive in those follicles. Weak immunostainings for IgG1 and IgM could be observed inside these follicles, and that for IgM was weak in the interstitium and theca layers (Fig. 28.2). Similar immunostaining of albumin, IgG1, and IgM was observed in antral follicles, where more layers of follicular cells and the antrum appear (Fig. 28.2). By contrast, immunostaining for the three serum proteins was all clear in the ovulated follicles [14], consistent with previous reports [11, 12].
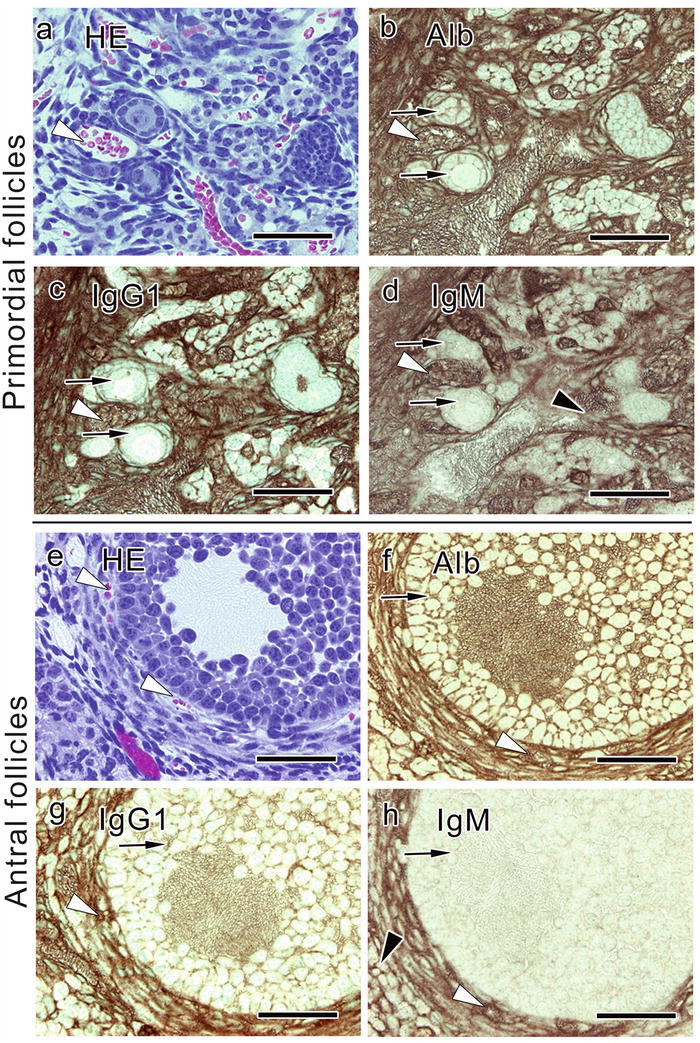
Fig. 28.2
Serial paraffin sections showing primordial (a–d) and antral follicles (e–h) obtained from living mouse ovaries with in vivo cryotechnique and stained with hematoxylin-eosin (HE, a, e) or immunostained for albumin (Alb, b, f), immunoglobulin G 1 (IgG1 , c, g), or IgM (d, h). The sections demonstrate that the strong immunoreactivities of all three serum protein s are observed inside blood vessel s (a–h, white arrowheads). The immunoreactivity of IgG1 and IgM is not clearly seen inside the follicles (c, d, g, h, black arrows), and the IgM immunostaining is weak also in the theca layers and interstitium (d, h, black arrowheads), as in or around the late primary follicles. The primordial follicles (a, b, c, d, black arrows) are composed of a single layer of flat follicular cells surrounding the oocyte, and antral follicles have more layers of follicular cells and antrum among them. Bars, 50 μm (The figure was modified from Zhou et al. [14])
Previous studies suggested that BFB substantially affects distributions of medium-sized serum molecules [11]. Indeed, immunostaining for IαI (220 kDa) or fibrinogen (340 kDa) observed with IVCT was weaker inside the ovarian follicles [14].
Double immunofluorescence labeling of basement membrane s and IgGfc or IgM in living mouse ovaries prepared by IVCT revealed that immunoreactivity of IgGfc was observed in the theca layers and interstitium , but weak inside ovarian follicles surrounded by the basement membranes which bordered the changes of IgGfc immunoreactivity [14]. IgM immunoreactivity was detected exclusively in blood vessel s around the theca interna and not in the ovarian follicles and interstitium. The immunolocalization of IgM was mostly restricted inside endothelium of the blood vessels surrounded by basement membranes.
Selective functions of molecular permeation throughout the follicular development until ovulation appear to be determined by BFB. In living mouse ovaries, the endothelial layers of blood vessel s determine in vivo permeability of large molecular weight proteins such as IgM , while the follicular basement membrane s are important for permselectivity of middle-sized molecules, such as IgG . Endothelium of blood vessels generally controls the passage of soluble component s and circulating blood cells into tissue parenchyma. Adhesion and communication between adjacent endothelial cell s are mediated by their intercellular junctions [16, 17]. Previous studies demonstrated that the tight junction s were critical for endothelial permeability [18, 19]. In living mouse ovaries, the functions of intercellular communication between endothelial cells are elusive regarding permeability of soluble serum protein s through blood vessel walls. Studies at an electron microscopic level using IVCT would be important to elucidate the components and structures responsible for intercellular communication between endothelial cells in blood vessels of ovaries and regulation of permselectivity against larger molecules.
The blood vessel s are surrounded by basement membrane s which function as a porous molecular sieve . The charge and structural arrangement of its components, including type IV collagen and heparan sulfate proteoglycans, are responsible for the molecular selectivity of the basement membranes [20, 21]. Leakage of large serum protein s out of blood vessels is prevented by basement membranes around blood vessels in some organs, such as kidneys [22, 23]. Follicular basement membranes in mouse ovaries would also have functions related to a molecular sieve of ovarian follicles and might be involved in regulation of developmental metabolic exchanges between inside and outside of the ovarian follicles. The permselectivity of basement membranes appeared different around follicles and blood vessels. Their functional difference might be explained by the difference in their components. During the follicular development and ovulation, the follicular basement membrane has to be dramatically changed [24, 25]. But significant relationship between basement membranes and the BFB, which are continuously sealing the ovarian follicles during all development stages, is supported by the observation that BFB was functioning at an early stage of folliculogenesis [26].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


