Chapter 4 Hierarchical organisation in the nervous system


The functional arrangement of the nervous system is hierarchical and can be likened, in some ways, to corporate structure (Fig. 4.1, Table 4.1). There are workers and management systems. The workers are those nerves that interact directly with the sensory receptors in the body and with the effectors (muscle). Thus the majority of the ‘worker’ nerves are found in the PNS. Conversely, management is confined to the CNS. It receives and processes sensory inputs, integrates information, and directs the worker outputs. There are a number of levels of management: junior, senior and executive. These levels are located to different extents in the spinal cord, brainstem and forebrain. There are also maintenance systems that keep the basal level of activity going in the body. These are represented largely by the autonomic nervous system and include visceral reflexes. For example, stimulation of baroreceptors results in cardiovascular changes to maintain appropriate blood pressure.
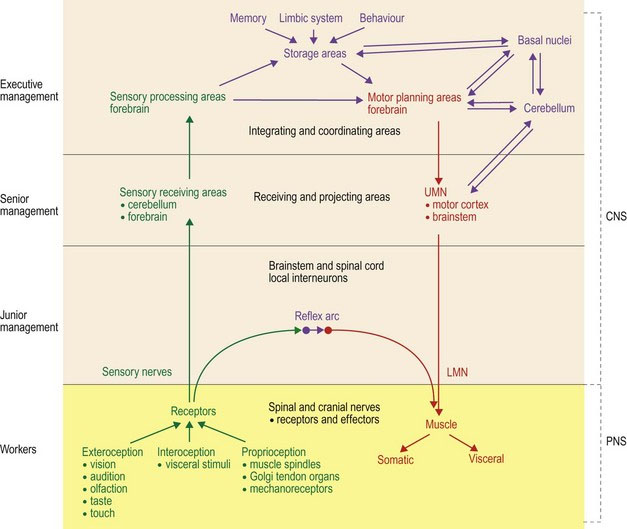
Table 4.1 Hierarchical arrangement of the nervous system
| Hierarchical position | Location of sensory system | Location of motor system |
|---|---|---|
| Workers Link between the PNS target organs and the CNS Involved in reflex functions | Sensory receptors and sensory nerves, with input via spinal and cranial nerves | LMNs connecting via NMJ to muscle |
| Junior management Interneuronal circuitry which links between input from sensory fibres, or UMNs, to LMN Involved in reflex functions | Spinal cord for afferent spinal nerves Brainstem for sensory input from CN V, VIII, IX, X Forebrain for sensory input from CN I, II | Spinal cord for connecting to LMNs of spinal nerves Brainstem for connecting to LMNs of CN III–VII, IX–XII |
| Senior management | Primary sensory receiving areas Cerebellum – subconscious proprioception Forebrain – visual, auditory, somatosensory cortices and olfactory lobe | UMN nuclei of the brainstem and the motor cortex of the forebrain |
| Executive management | Diverse areas of the brain processing and interpreting sensory input. Interconnections between sensory association areas, storage (memory), behaviour and emotion centres and motor planning centres with links to the cerebellum and basal nuclei | |
Overview of motor and sensory systems
Key points





Overview of motor (output) systems
The functions of the motor system are (a) to maintain posture and provide a stable platform for movement, (b) voluntary movement and locomotion, and (c) visceral motor function (see Chapter 5).
Upper motor neurons, lower motor neurons and the neuromuscular junction
The motor system can be divided into two main components – upper motor neurons (UMNs) and lower motor neurons (LMNs). The UMNs are completely contained within the CNS, thus an alternative name for them could be ‘central motor neurons’. The UMN cell body is located in a motor nucleus of the brainstem or the motor cortex of the forebrain. Their axons connect to LMNs either by synapsing on them directly or indirectly, via interneurons. The LMNs have their cell body in the CNS (brainstem or spinal cord). Their axons project into the PNS, via cranial or spinal nerves, to connect with striated or smooth muscle at the neuromuscular junction; thus an alternative name for them could be ‘peripheral motor neurons’. Note that while the soma of the LMN connecting to striated muscle lies in the CNS, its long axon is located in the PNS. The ANS has two LMNs in series, with the cell body of the second LMN located in a peripheral ganglion (see Chapter 12). The UMNs, along with other management components of the motor system, organise and direct the activity of the LMN. When stimulated LMNs induce muscle contraction. Thus the UMNs are part of a motor management system, while the LMNs with their associated muscles are analogous to the workers.
The neuromuscular junction (NMJ) (see Fig. 1.4) connects the electrical activity of the LMN to the depolarisation of the muscle, using chemical messengers, such as acetylcholine (ACh). Even in the absence of action potentials, acetylcholine is continually being released at the NMJ causing miniature depolarisations of the muscle end plate. These miniature end plate potentials result in end plate noise that can be detected electromyographically using needle electrodes. Release of ACh also contributes trophic support to the muscle fibres, which when lost, as occurs when the LMN is destroyed, is associated with rapid, severe, neurogenic atrophy of the muscle fibres (see Chapter 5). When a volley of action potentials arrives at the motor end plate, large amounts of ACh are released and the muscle is stimulated to contract proportionally.
There are two basic activities of striated muscles in the body and limbs, extension and flexion of joints. Extensor muscles are antigravity muscles as they permit the animal to resist the effects of gravity. Extensor muscles increase the joint angle. For the thoracolumbar vertebral column, flexor (hypaxial) muscles work in conjunction with extensor (epaxial) muscles to resist the effects of gravity on the trunk. Antigravity muscle activity has to dominate flexor muscle activity, otherwise the animal would be recumbent. However, in the normal animal, extensor dominance is modulated by the action of inhibitory UMNs synapsing on the LMN supplying extensor muscles. Thus the overall descending UMN activity in the normal animal is inhibitory to antigravity extensor muscles. Damage to these inhibitory UMN pathways results in extensor dominance and spasticity (Fig. 4.2).
Worker and management systems
Sensory systems
Sensory input that is received in the primary sensory cortices is projected to nearby association areas where it is processed, ranked in terms of importance and, from there, integrated with other information. The association and integration areas are the equivalent of the executive management for the sensory systems. For example, visual input via cranial nerve II is received in the visual cortex in the occipital lobes. The executive system for visual input will process that information in the nearby visual association area and link it with other processed sensory inputs (e.g. touch, audition), stored information (memory), emotion-generating areas of the brain (limbic system) and information from other integrating and coordinating centres such as the cerebellum and basal nuclei. The locations of the main functional regions of the cortex are comparable in all mammals, but their extent varies: for example in the carnivore the primary sensory and motor areas and olfactory cortex comprise 80% of the cortical surface whereas in primates the same zones make up less than 20%. The relationship is reversed for the associative cortex in these two groups of animals, while rodents and lagomorphs have almost no associative functions at all (see Fig. 4.16).
The workers and maintenance crew: The reflex arc

A reflex arc involves both the PNS and the CNS. It has a sensory/afferent nerve in the PNS that brings information into the CNS where the input is linked to a motor/efferent nerve that connects back to a muscle (Fig. 4.3). Via that linkage, the stimulus of the sensory nerve elicits a stereotypical motor output and muscle activation. The neural wiring of a reflex arc is laid down during embryological development. Thus, unlike a learned motor response, its function does not depend on experience. An example of a learned motor activity is the menace response in which the animal blinks in response to a menacing gesture made to the eye. By comparison, the palpebral reflex, in which the animal blinks in response to tactile stimulation of the palpebrum, is present from birth.
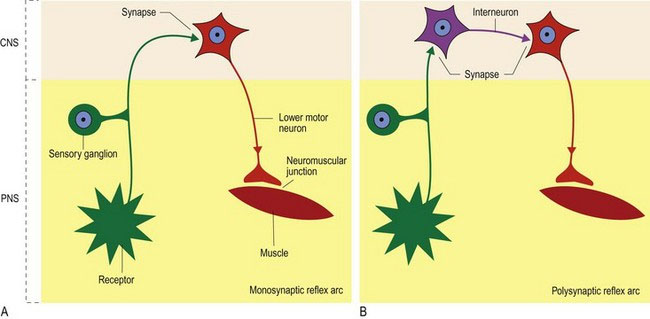
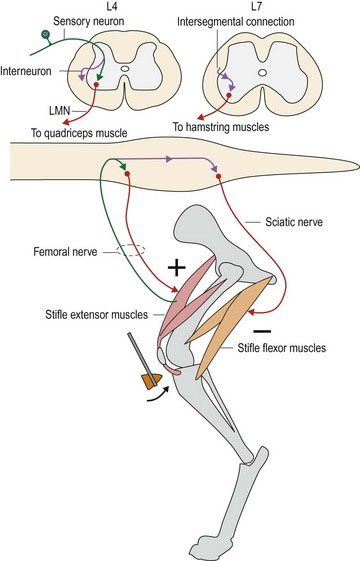
A typical reflex is the patellar, or ‘knee jerk’ reflex, in which tapping the patellar ligament with a plexor (patellar hammer), causes reflex contraction of the quadriceps muscle and stifle extension (Fig. 4.4). Simultaneously, the antagonistic semitendinosus and semimembranosus muscles are reflexively inhibited from contracting. Sensory input is via the femoral nerve. In dogs, this nerve synapses directly onto the LMNs in the ventral horns of L4–L6 spinal cord segments, which supply the quadriceps muscle, also via the femoral nerve. Note: a monosynaptic reflex like this is unusual but may include stretch/myotatic reflexes (see Chapter 5); interneurons are usually interspersed between afferent and efferent neurons, resulting in a polysynaptic reflex. Simultaneously, the sensory input also travels caudally, via interneurons, to spinal cord segments L6–S1 to synapse on interneurons that inhibit LMN-induced contraction of the hamstring muscles. Thus, contraction of the quadriceps muscle is unopposed.
Junior management: Spinal cord and brainstem
Key points
 Nerve fibres in the spinal cord enable local connections between sensory and motor nerves for reflex activity. They also provide links between the body and the brain for sensory and motor functions.
Nerve fibres in the spinal cord enable local connections between sensory and motor nerves for reflex activity. They also provide links between the body and the brain for sensory and motor functions.
 Both the grey and white matter can be divided functionally into distinct regions.
Both the grey and white matter can be divided functionally into distinct regions.



 UMN tracts in the spinal cord originate in the brain and mainly influence γ-LMN, via interneurons.
UMN tracts in the spinal cord originate in the brain and mainly influence γ-LMN, via interneurons.

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


