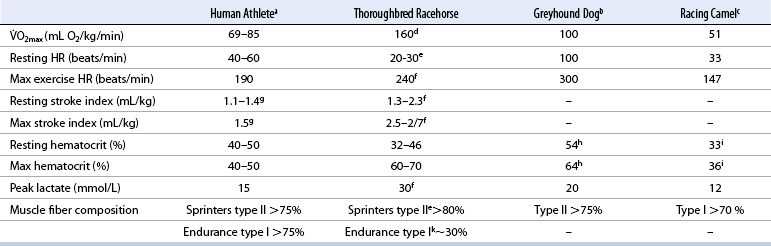
CHAPTER 2 Of all the athletic species in the world, four have dominated exercise physiology studies. These are (1) the athletic human, (2) the racehorse, (3) the greyhound, and (4) the racing camel (Figure 2-1). Of these species, the human athlete has received the most attention and the greyhound and the camel the least. The racehorse occupies an intermediate position. Interest in the physiologic changes that occur during exercise in humans began at the end of the nineteenth century. By the early 1920s, scientists began to address practical questions, including the possible biochemical and physiologic causes of the fatigue that develops during exercise (Gordon, 1925; Hill, 1923). However, it was only from the late 1960s that research in sports medicine and the exercise sciences began to develop as a reputable academic discipline. Perhaps two key reasons stimulating this development were (1) the trend toward an increased interest in health internationally, and (2) the growing dominance of international sport by athletes from eastern European countries, especially the former German Democratic Republic (East Germany). The success of athletes from socialist countries posed a challenge to the Western nations, especially those from the United States, who were anxious that the perceived superiority of the capitalist system in all spheres of human endeavor should not be undermined. Thus, financial and political support for sports-related research increased in most Western countries, stimulating the rebirth of these disciplines on a global scale. In contrast, the exercise sciences have received little financial and intellectual support in the horse racing community. In his book, The Fit Racehorse, Tom Ivers (1983) has written that “[the racehorse] industry honors the past more ferociously than it defends its own existence.” This ethos likely remains true in the twenty-first century. It is very apparent that the principal focus of the horse racing industry remains in its past, that is, in examining the breeding history of the species, especially of its champions. Limited attention is paid to its future, which should involve the application of scientific knowledge and techniques to the study of horses and horse racing. As a direct result, progress in equine exercise science research lags behind human exercise research science by decades. A second possible explanation is that in the majority of races, the winning time is unimportant because there are no additional financial rewards for record performances. Patrick Cunningham (1991) wrote that “nobody is much interested in improving the average racing times . . . what does it matter if all horses race 10 percent faster?” Hence there is no incentive to improve the performances of all racehorses progressively and in a systematic manner. Competitive racing for humans and horses shares a common origin—the use of these species for transport. The Greeks and the Romans used runners to deliver messages by foot, often over long distances. This tradition continued in Britain in the tenth century a.d. However, only from the seventeenth century onward was running established as a competitive sport, originating mainly in Britain (Noakes, 1992). The very earliest human footraces were usually over extremely long distances. It is recorded that a race of 237 kilometers (km) (147 miles) took place in Rome during the Roman Empire (Noakes, 1992). Recognized distances for modern footraces range from sprints of 100 meters (m) to ultramarathon races of 1000 km or more, lasting many days. The top speeds achieved by human sprinters exceed 36 kilometers per hour (km/h; 22 miles/h), whereas speeds of around 16 km/h (10 miles/h) are more common in marathon races of up to 100 km and of 6 to 8 km/h (4 to 5 miles/h) in races of 1000 km or more. Horse racing originated with the Bedouins of the Middle East, who dehydrated their horses and trained them to race to the nearest water hole. It was an event in the Greek Olympic Games as early as 664 b.c. The first recorded race meeting in Britain was in the twelfth century in London. However, it was only in the late seventeenth century (circa 1665) that organized Thoroughbred horse racing, as we now know it, first took place in New York and in Newmarket, England (Kidd, 1976). The camel is known for its endurance ability in hot, dry environments. In the Middle East, specially bred camels are raced over distances of 4 to 10 km and achieve speeds of approximately 36 km/h (22 miles/h) (Evans et al., 1992). From as early as 1835, greyhounds were raced against each other, sometimes in races involving as many as 64 participants (Genders, 1990). In 1858, the sport was officially organized with the formation of a governing body, the National Coursing Club, in Britain. The endurance ability and resistance to cold temperatures of the husky and related species have long been recognized. These dogs are the principal means of transport for polar travel. The first races were held in Alaska in 1907 over 600 km (400 miles) (Sayer, 1989). The most famous modern race is the Iditarod, in which dogs race from Fairbanks to Nome in Alaska over 1050 to 1150 km. This race takes 8 to 14 days to complete (www.iditarod.com/about/, 2011). In humans, there is a trend for the best athletes to have the highest Resting heart rate (HR) values are in the low twenties in fit horses, whereas values of 40 to 60 are more usual in athletic humans under the same conditions. During exercise, maximal HR values of between 240 and 250 beats per minute (beats/min) have been recorded in racehorses, whereas maximal values in the range of 180 to 200 beats/min are more common in athletic humans (Evans and Rose, 1988; Noakes, 1992). The greyhound has a maximal heart rate of about 300 beats/min, only a threefold increase from resting values (Snow, 1985). The camel, in contrast, has the lowest TABLE 2–1 Data from aNoakes; bSnow; cRose and colleagues; dRose and colleagues; eSnow and Vogel; fPhysick-Sheard; gGanong; hSnow and colleagues; iEvans and colleagues; jMcMiken; kRose. The resting stroke index (SI, divided by body weight) for horses is between 1.3 and 2.3 mL/kg, increasing to 2.5 to 2.7 mL/kg during maximal exercise (Physick-Sheard, 1985), similar to the human resting value of 1.1 to 1.4 mL/kg, which increases to around 1.5 mL/kg during maximal exercise (Ganong, 1985). Although the stroke index increases in both humans and horses, the increase is, at most, of the order of one- to twofold. Hence it is the much larger (fourfold to 10-fold) increase in HR that is the main contributor to the increase in CO during exercise in both racehorses and human athletes. The hematocrit is the percentage of the total blood volume occupied by red blood cells (RBCs). The human athlete maintains this hematocrit value at between 40% and 50% (Wilmore and Costill, 1988). During exercise, in humans, the hematocrit tends to rise slightly as a result of a fall in the amount of fluid, the plasma volume, in which the RBCs circulate. The total number of RBCs contained in that volume may increase only slightly during exercise in humans. In contrast, the horse has the unique ability, specifically during exercise, to release a large number of RBCs from the spleen. As a result, the hematocrit of the horse can increase from around 32% to 46% to 60% to 70% during maximal exercise (Snow and Vogel, 1987). This ability dramatically increases the oxygen-carrying capacity of the horse’s blood during exercise. Greyhounds have high resting hematocrit levels of about 54%; these increase to around 64% during maximal exercise. It is not known whether this is caused by the release of RBCs from the splenic reserve or results from a decrease in plasma volume causing an increase in the concentration of red blood cells (Snow et al., 1988). The hematocrit of the racing camel increases from 33% at rest to 36% at maximal exercise (Evans et al., 1992). Of the four athletic species, the camel, therefore, has the lowest concentration of RBCs. This, together with a relatively low maximum HR, might explain the relatively low Some human athletes have attempted to mimic this physiologic response by using an illegal technique known as “blood doping.” In this procedure, RBCs are withdrawn from the athlete and frozen. At some point, usually hours before competition, the stored RBCs are reinjected, thereby increasing the oxygen-carrying capacity of the blood and potentially aiding performance (Buick, 1980). This procedure appears to have limited effects in the horse, possibly because of the innate capacity of the horse to increase the packed cell volume (PCV) in response to exercise. Another way to increase this effect is by using high altitude in training programs with the current recommendation of live high, train low for altitude training in people. However, in both humans as well as horses, the effects are relatively small and not consistent (de Paula and Niebauer, 2010; Wickler and Anderson, 2000). The maximum arteriovenous oxygen difference, or (a-v)DO2, measured in the horse during maximal exercise is only very slightly greater than values measured in elite human athletes under similar conditions. Thus, a larger (a-v)DO2 accounts for only about 23% of the greater Although Two important factors are locomotive efficiency, which is the oxygen cost per kilogram per kilometer traveled, and the percentage of An important factor in determining success in human runners and cyclists is economy of locomotion, or a lower than average oxygen cost at any running or cycling speed (Coyle et al., 1991; Noakes, 1992). This has yet to be evaluated in racehorses and the other athletic species. However, when comparing the oxygen cost of exercise in camels and horses, it is clear that the oxygen cost per kilometer traveled in camels is much less than in the horse, indicating superior economy of locomotion. Thus, the greater capacity for oxygen transport in the racehorse is the result of a larger capacity to increase CO, with heart rate being the main contributor; the greater oxygen-carrying capacity of the blood during exercise; and finally, a small increase in the capacity to extract oxygen in the active muscles, measured as a greater (a-v)DO2 (Evans and Rose, 1988).
Comparative aspects of exercise physiology
Main athletic species
The human athlete
Distances
The equine athlete
The racing camel
Racing dogs: The greyhound
Racing dogs: The husky
Comparative physiology of the athletic species
Oxygen consumption
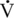 O2max values. Elite human athletes have
O2max values. Elite human athletes have 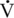 O2max values ranging between 69 and 85 milliliters oxygen per kilogram per minute (mL O2/kg/min), whereas Thoroughbred racehorses have
O2max values ranging between 69 and 85 milliliters oxygen per kilogram per minute (mL O2/kg/min), whereas Thoroughbred racehorses have 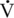 O2max values twice as high, about 160 to 200 mL O2/kg/min (Noakes, 1992; Rose et al., 1988).
O2max values twice as high, about 160 to 200 mL O2/kg/min (Noakes, 1992; Rose et al., 1988).
Cardiac output
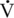 O2max in this species is in excess of 100 mL O2/kg/min, although this has proven difficult to measure.
O2max in this species is in excess of 100 mL O2/kg/min, although this has proven difficult to measure.
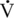 O2max value (51 mL/kg/min) of the four common athletic species, the horse, the greyhound, the camel, and the human (Evans et al., 1992) (Table 2-1). The racing camel can increase its HR fourfold from a resting rate of about 33 beats/min to about 150 beats/min during maximal exercise.
O2max value (51 mL/kg/min) of the four common athletic species, the horse, the greyhound, the camel, and the human (Evans et al., 1992) (Table 2-1). The racing camel can increase its HR fourfold from a resting rate of about 33 beats/min to about 150 beats/min during maximal exercise.
Stroke index
Blood oxygen content
 O2max of this species.
O2max of this species.
 O2max of the racehorse compared with the elite human athlete; the much greater CO and oxygen-carrying capacity of the blood accounts for the other 77% (Physick-Sheard, 1985).
O2max of the racehorse compared with the elite human athlete; the much greater CO and oxygen-carrying capacity of the blood accounts for the other 77% (Physick-Sheard, 1985).
 O2max as a predictor of athletic ability
O2max as a predictor of athletic ability
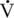 O2max is generally considered the best predictor of athletic potential, there is evidence to dispute this belief (Noakes, 1988). For example, although elite human athletes do have high
O2max is generally considered the best predictor of athletic potential, there is evidence to dispute this belief (Noakes, 1988). For example, although elite human athletes do have high 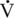 O2max values, so, too, do many less conditioned athletes. Athletes with similar athletic abilities may have quite different
O2max values, so, too, do many less conditioned athletes. Athletes with similar athletic abilities may have quite different 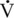 O2max values (Noakes, 1992).
O2max values (Noakes, 1992).
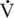 O2max that can be sustained during prolonged exercise (Coetzer et al., 1993; Hammond et al., 1984; Noakes, 1992). Subjects able to run at a higher percentage of their
O2max that can be sustained during prolonged exercise (Coetzer et al., 1993; Hammond et al., 1984; Noakes, 1992). Subjects able to run at a higher percentage of their 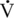 O2max for longer periods exhibit superior fatigue resistance. For example, the
O2max for longer periods exhibit superior fatigue resistance. For example, the 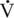 O2max of the Thoroughbred racehorse is about three times greater than that of the racing camel, which is of similar mass (Rose et al., 1992) (see Table 2-1). Yet the camel can exercise at a very high intensity (100%
O2max of the Thoroughbred racehorse is about three times greater than that of the racing camel, which is of similar mass (Rose et al., 1992) (see Table 2-1). Yet the camel can exercise at a very high intensity (100% 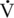 O2max) for a much longer period than can the horse (18 min versus 3–5 min). Thus, the performance of the camel in races lasting more than 10 to 15 minutes is likely to be more similar to that of the racehorse because of the former’s ability to exercise at a much higher percentage of
O2max) for a much longer period than can the horse (18 min versus 3–5 min). Thus, the performance of the camel in races lasting more than 10 to 15 minutes is likely to be more similar to that of the racehorse because of the former’s ability to exercise at a much higher percentage of 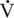 O2max for much longer despite a substantially lower
O2max for much longer despite a substantially lower 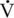 O2max. Hence, in the assessment of an athlete’s potential, it is necessary to consider not only the
O2max. Hence, in the assessment of an athlete’s potential, it is necessary to consider not only the 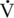 O2max but also the percentage of
O2max but also the percentage of 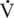 O2max that can be sustained during prolonged exercise.
O2max that can be sustained during prolonged exercise.
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Comparative aspects of exercise physiology

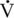 o2max
o2max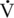 O2max) is the maximal amount of oxygen used by the athlete during maximal exercise to exhaustion.
O2max) is the maximal amount of oxygen used by the athlete during maximal exercise to exhaustion. 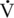 O2max is sometimes referred to as the peak aerobic power.
O2max is sometimes referred to as the peak aerobic power.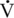 O2max of the Thoroughbred racehorse compared with the elite human athlete.
O2max of the Thoroughbred racehorse compared with the elite human athlete.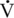 O2max, Heart Rate, Peak Blood Lactate, Hematocrit, Stroke Index, and Muscle Fiber Composition
O2max, Heart Rate, Peak Blood Lactate, Hematocrit, Stroke Index, and Muscle Fiber Composition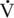 O2max values of Standardbred (165–180 mL/kg/min) and Thoroughbred, (164–200 mL/kg/min) horses. Clearly, if
O2max values of Standardbred (165–180 mL/kg/min) and Thoroughbred, (164–200 mL/kg/min) horses. Clearly, if 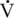 O2max was the sole predictor of athletic ability, the value should be much higher in Thoroughbred than in Standardbred horses. Hence factors other than
O2max was the sole predictor of athletic ability, the value should be much higher in Thoroughbred than in Standardbred horses. Hence factors other than 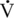 O2max must be important in determining the superior athletic ability of the Thoroughbred racehorse.
O2max must be important in determining the superior athletic ability of the Thoroughbred racehorse.Only gold members can continue reading. Log In or Register to continue
