Fig. 1.
Ground reaction forces produced by rats moving along a runway. (a) Rats are conditioned to move repeatedly along a runway in which three force platforms (plates 1–3) are embedded in the ground surface. Ground reaction forces (forces exerted through the limbs onto the ground) are recorded in three orthogonal directions: vertical (red arrow), fore-aft (blue arrows) and lateral (green arrows). The rat contacts each platform first with a forelimb followed by the ipsilateral hindlimb, so that forces from the right and left limbs can be recorded separately (right and left in (b–d)). (b) Peak vertical forces are approximately equal for fore- and hindlimbs, although impulse (area under the curve, indicated in light blue) differs between fore- and hindlimbs. (c) Forelimbs generate most of the braking forces while the hindlimbs generate more propulsion. (d) Lateral forces are small for both fore- and hindlimbs. For each graph, solid lines represent mean data from unlesioned animals (n = 12), thin lines are ±standard error.
Our laboratory uses custom-built force platforms (41). Three platforms, each measuring approximately 11 cm × 11 cm, are positioned level with the surface of the runway (180 cm × 15 cm) and arranged in series. The size of the plate surface is important. If the plates are too small, the rat will miss contact with a plate as it passes over. If the plates are too large, more than one limb will contact the plate at one time, which does not allow the investigator to distinguish which limb is responsible for the forces measured. Each platform measures force in three orthogonal directions (vertical, fore-aft and medio-lateral). The position of the limb on each platform can be calculated from the ratio of the input from vertical force sensors located at each corner of the platform (41). Outputs from the platforms are amplified, digitally sampled (1,000 Hz) and acquired simultaneously with digital video recordings (Midas, Xcitex Inc, Boston, MA). A digital camera (125 Hz, Motionscope 1050; Redlake MASD Inc.) is positioned perpendicular to the runway. An infrared beam positioned across the runway ahead of the series of three force platforms triggers simultaneous GRF and digital video recordings each time the rat approaches the platform. A second beam positioned on far end of the platforms stops data recording and an LED display indicates the time taken for the rat to traverse between the beams, providing a measure of average velocity over the platforms.
Data acquired from our custom-built force platforms is analyzed using custom-written software, although commercially available force platforms will have accompanying software. Data for each pass is digitally filtered using a fourth order Butterworth Filter (cut-off 40 Hz). For each pass, the force data from each plate are matched with the identity of the limb that has contacted the plate, using time-synchronized digital video recording (Fig. 1a and Sect. 3.1.3). Data for each limb from different passes (ten passes minimum) across the plates are averaged to obtain a representative force for each limb. In order to compare performance between individuals and between groups of animals, we examine key features of the GRF profile for each limb such as peak force (e.g., maximum vertical, maximum braking and maximum propulsive forces) and impulse (e.g., area under the force vs. time graph for vertical, braking and propulsive forces) (Fig. 1b, c). The value of these variables are determined for each pass and averaged for each limb of an individual animal.
3.1.2 The Importance of Locomotor Velocity
In order to increase the chances of acquiring data when the rats are moving at a steady speed across all three platforms, the rats are conditioned to repeatedly traverse back and forth along the runway for a food reward. A steady speed is necessary because ground reaction forces are only consistent when net forward acceleration is 0, since animals use their limbs differently if they are accelerating or decelerating. Rats must be motivated to shuttle to and fro but at the same time must be relaxed enough so that they are not dashing from one end of the runway to the other. We achieve this by conditioning the animals over several weeks to expect a food reward (a Cheerio) as they reach either end of the runway, so that they shuttle calmly back and forth across the platforms.
In addition to ground reaction forces, many other locomotor parameters can be rapidly and accurately obtained from the force plates. The plates record the position and time of contact of each paw as the limb is placed on the platform, so that spatial and temporal variables can be obtained for individual limbs (stride lengths, stance time) and as well as for interlimb coordination (timing overlap) (22).
An important consideration for measurement of all locomotor parameters, including ground reaction forces, is the speed with which the animal is travelling. Stance time, stride lengths, peak forces are just a few variables which differ significantly at different speeds. It is important to note that, after injury, animals will often move more slowly than prior to injury and many locomotor parameters change simply because of the animal’s reduced velocity. For valid comparisons of locomotor parameters, it is essential that pre- and post-surgical data be collected at the same speed. Because we cannot control the speed with which animals move overground unrestrained (unlike with treadmill locomotion, for example), we record many passes during pre-surgical data collection. We record the speed with which the animal moves for each pass and then, during analysis, we group the data for each individual according to speed. This ensures that there will be pre-surgical data sampled at a number of different velocities, allowing us to match pre-surgical data to post-surgical data at comparable speeds.
3.1.3 Kinematic Data Collection
Digital video recording of rats as they move over the force platforms allows the identification of the limbs which contact each platform (e.g., right or left, forelimb or hindlimb), information which is necessary for the correct interpretation of GRF data. Using custom-written software, the data from force plates will also provide limb timing, step distances and limb overlap measurements using the onset and off-set timing of individual limbs on all three force platforms (22). This information provides very precise timing and distance measurements because sampling rate of the GRF data (1,000 Hz) is much higher than that of most digital video data (e.g., 25–60 Hz), the more common method with which to obtain limb timing and step distance information.
We also use digital video to record the position and orientation of the limb segments in each video frame as the animal moves through the stride cycle. To identify the position of limb segments in each video frame, reflective markers are placed at the ends of each segment prior to recording. Markers (2-mm diameter) can be fashioned from reflective tape (3M, St. Paul, MN, USA) shaped into a shallow cone and glued to the ends of each limb segment or over joints which have been clipped free of fur. During analysis, the markers are identified in each video-frame, allowing computer-assisted digitization of segment positions throughout the stride (ProAnalyst; Xcitex Inc., Boston, MA, USA). This information is then used to construct a model of each limb as a series of linked segments (Fig. 2). The kinematic data obtained from this analysis (e.g., limb segment positions and joint angle changes throughout the stride) are used to quantitatively describe the action of each limb during locomotion (42–45).
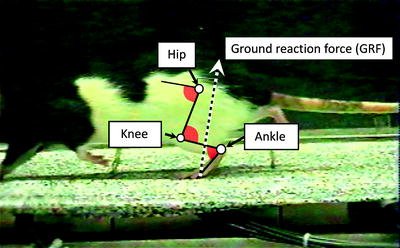
Fig. 2.
Video image of rat trotting over force platforms as it moves past the camera. Computer-assisted digitization of reflective markers placed on the limb segments allows the estimation of joint positions (hip, knee, ankle) and segment locations in each video frame. Reconstruction of the limb as a series of linked segments allows calculation of joint angles (red shading) throughout the stride cycle. Simultaneous collection of video data and ground reaction forces allows the calculation of joint torques and powers by synchronizing limb segment positions in each video frame with the magnitude and position of the ground reaction force vector (dashed line) obtained from force plate measurements.
3.1.4 Mechanical Analysis of Limb Movement
The simultaneous collection of GRF data and kinematic data allows incorporation of kinetic information into the linked segment model of the limb described above, thus providing information about the mechanical action of the limbs (40). Joint torques and joint power are two commonly determined measures, which can be obtained from kinetic and kinematic data using a method called inverse dynamic analysis (40). This method is commonly used for human gait analysis, and has proven valuable for the diagnosis of pathological gait. Biomechanical analyses have been investigated only recently in laboratory rodents (46, 47). In our laboratory, we have used the inverse dynamic method to examine joint torques and joint power for the joints of all four limbs after peripheral nerve injury (Bennett et al., unpublished data). After loss of ankle extensor innervation, we found that rats alter the power at the joints of the lesioned hindlimb in complex ways, and additionally increase the net mechanical power produced by the unlesioned limbs to compensate for the reduced power of the lesioned limb. Thus, inverse dynamic analysis can provide quantitative biomechanical information on how rats use their limbs before and after injury.
3.2 Skilled Locomotion
Skilled locomotion refers to locomotor tasks that require the rat to move over challenging terrain. Rats are naturally nimble and are able to negotiate many obstacles and structures. In our laboratory, we examine their ability to move across a horizontal ladder for a food reward (11, 22–24). Other researchers have similarly examined locomotion over ladders, grids or along beams of different or tapering widths (1, 13, 48–50). We prefer to examine skilled locomotion over a horizontal ladder rather than a grid because the wire ladder rungs present a consistent surface for each paw. A grid, with its support bars running in two dimensions, can present a slightly different surface for each paw, depending on the position of the paw relatively to the grid bars. The beam is similar to the ladder in that it presents a consistent surface for each paw placement, although the texture of the beam surface will have an effect on the amount of friction afforded to each paw.
Similar to conditioning for the collection of kinetic and kinematic data (Sect. 2.1 above), we condition rats to shuttle repeated back and forth across a horizontal ladder for a food reward as they reach either end (Fig. 3a). The ladder apparatus is a 124-cm long runway, in which the central 80 cm portion of the floor of the runway consists of a horizontal ladder (2 mm diameter wire rungs placed at 2-cm intervals (24)). Rats are considered sufficiently conditioned when they cross the ladder willingly up to ten times in succession. Consistent with our measurement of overground locomotion (Sect. 3.1 above) and skilled forelimb use (Sect. 4.1 below), we therefore examine the effect of our experimental manipulations on the performance of well-known ladder task (see Sect. 2.2.3). Rats are digitally videotaped from a lateral perspective as they repeatedly cross the ladder. A mirror angled at 45° beneath the ladder allows both the lateral and ventral views to be recorded in each video frame (Fig. 3a).
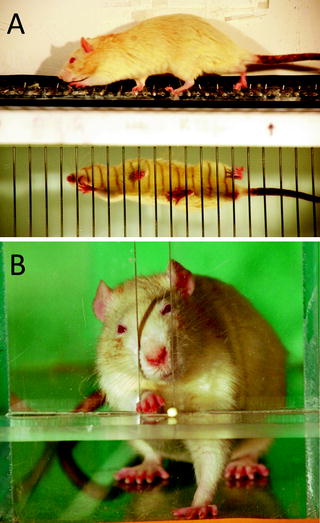
Fig. 3.
(a) Video images of rats crossing a horizontal ladder. A mirror placed below the ladder and positioned at 45° to the horizontal allows lateral and ventral views to be recorded simultaneously. Rats are conditioned to repeatedly shuttle back and forth across the ladder so that multiple independent strides can be recorded so as to obtain a representative sample of ladder performance. (b) Video images of rats reaching for a food pellet from a shelf located outside the reaching box. Rats are conditioned to retrieve a food reward at rear of the reaching box after each reaching attempt, to ensure that each reach is independent of the previous attempt.
During analysis of digital video, data is collected from only one stride per pass to ensure independence of different samples from the same animal (Sect. 2.2.2). We collect ten samples from each animal to obtain a representative measure of each animal’s performance. In keeping with the use of quantitative measures, we assess whether each paw made a correct step (full palmar or plantar contact with the rung maintained throughout weight support), an abnormal step (e.g., wrist only) or an error (e.g., paw contact with a rung was missed or lost, resulting in loss of equilibrium). For each paw, these three events are counted over the ten strides, providing quantitative measurement along with an indication regarding the quality of paw placement.
4 Assessing Forelimb Use
Rats have a natural ability to use their forelimbs for many behaviours in addition to locomotion, including exploration and manipulation of objects, particularly food. While there are many tests available to assess forelimb function (1, 2), our laboratory has focused on the pellet retrieval task (see Karl and Whishaw, Volume I, ch. 6, and below), and the paw preference task. Originally developed to assess forelimb function in experimental brain lesions (51), these tests are thought to be analogous to assessments of arm and hand function in humans (52). Tests of forelimb function have also proven useful for assessing the effects of cervical SCI in rat models (23, 24, 53–55).
4.1 Skilled Reaching
Although rats are skilled at using their paws to handle food, they nevertheless need to be conditioned to use their forepaws to prehend food, a task they normally accomplish with their mouths. The Whishaw skilled pellet retrieval task (Karl and Whishaw, Volume I, ch. 6) is a well-described task which can be used to assess forelimb function under many experimental conditions. Our use of the task varies little from the original descriptions of this test (56, 57). Rats are placed in a rectangular box (45 cm × 12 cm) with a ledge attached to the outside of the box. A flavoured sugar pellet (4.0 mm × 3.3 mm, 45 g, BioServe, Inc) is placed into a shallow depression on the ledge. The rat can access the pellet through a vertical opening in the wall of the box which is wide enough for a paw to fit through but not the rat’s snout. With repeated attempts, rats learn to use their forepaw to reach the pellet and with further exposure will learn to grasp the pellet with the digits to successfully retrieve it (Fig. 3b).
As for many behavioural tasks, there are a variety of methods with which to score performance. In our laboratory, we count the number of successful attempts out of 20 possible reaches. A successful reach is one in which the pellet is grasped with the digits, brought to the mouth and eaten in one attempt. If the rat misses the pellet on the first attempt, or drops it before it reaches its mouth, this is not considered a successful reach. Other studies have used different measures to assess reaching success, including counts of the number of pellets successfully retrieved in a fixed amount of time, or the number of attempts used to retrieve a single pellet (2, 58). These endpoint measures are useful because they provide a quantitative score against which performance can be compared before and after experimental SCI or therapy. Nevertheless, endpoint measures do not provide any information regarding how the animal is performing the task (2). After injury, rats frequently use a variety of “abnormal” movements to reach and retrieve pellets and similar endpoint scores can be achieved using different movement strategies (32). To address this, a more detailed analysis of reaching behaviour is required (57, 59). The reaching action of the forelimb can be divided into ten sequential components, and frame-by-frame analysis of videotape recordings allows the scoring of the presence or absence of each component (57, 59). We have used this method to examine the reaching movements used by rats after combined pyramidal tract and spinal cord injury (23, 24).
< div class='tao-gold-member'>
Only gold members can continue reading. Log In or Register to continue
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


