Chapter 6 Ascending somatic sensory tracts and conscious sensory systems
Key points
 Sensory information is received from exteroceptors, interoceptors and proprioceptors and is projected to specific regions of the forebrain for conscious perception. Proprioceptive input is also projected to the cerebellum for subconscious processing. Most sensory inputs are also projected to the brainstem ascending reticular activating system for arousal (see Chapter 11).
Sensory information is received from exteroceptors, interoceptors and proprioceptors and is projected to specific regions of the forebrain for conscious perception. Proprioceptive input is also projected to the cerebellum for subconscious processing. Most sensory inputs are also projected to the brainstem ascending reticular activating system for arousal (see Chapter 11).

 General proprioception is the sense of the relative positions of body parts.
General proprioception is the sense of the relative positions of body parts.


Receptors
Exteroceptors are activated by stimuli in the immediate external environment such as temperature and touch. They include both encapsulated (Pacinian corpuscles and Ruffini endings) and free nerve endings that respond to pressure, vibration and distortion of the tissue. They may adapt to a constant stimulus either rapidly or slowly. Exteroceptors also include those receptors concerned with special sensations, such as audition, vision and gustation (taste) (see Chapter 10).
Pathway: Most conscious sensory systems share a common format that comprises a three-stage system:
1. A receptor and an axon with a cell body in the ganglion in the PNS;
2. A relay section in the CNS comprising a central pathway that has one, or more, synapses in specific nuclei. It eventually joins, or runs adjacent to, a pathway called the medial lemniscus through the brainstem to the thalamus;
3. Thalamocortical projection to the somatosensory cortex of the cerebrum.
Sensory modalities that do not fit this three-stage structure include olfaction, vision and some nociceptive pathways (Table 6.1).
Table 6.1 Ascending tracts, their functions and locations in the spinal cord
| Modality | Tract | Location |
|---|---|---|
| Nociception | Fasciculus gracilis and fasciculus cuneatus (skin) | DF |
| Spinocervicothalamic tract (skin) | LF | |
| Spinothalamic tract (skin, viscera and body) | LF | |
| Spinoreticular tract (viscera and body) | VF | |
| Spinomesencephalic tract | VF | |
| Temperature | Spinothalamic tract (viscera and body) | LF |
| Discriminative touch, pressure and conscious proprioception | Fasciculus cuneatus (from the body cranial to T8) | DF |
| Fasciculus gracilis and dorsal spinocerebellar tract (from the body caudal to T8) | DF | |
| Arousal and behaviour | Spinothalamic tract | LF |
| Spinoreticular tract | VF | |
| Spinomesencephalic tract | VF | |
| Subconscious proprioception and cerebellar input | Dorsal and ventral spinocerebellar tract (from the caudal half of body) | LF |
| Spinocuneocerebellar and cranial spinocerebellar tracts (from the cranial half of body) | DF and LF, respectively | |
| Spinoreticular tract | VF | |
| Spinopontine tract | VF | |
| Spinovestibular (from the neck) | VF | |
| Spino-olivary tract | VF |
DF = dorsal funiculus, LF = lateral funiculus, VF = ventral funiculus.
See also Fig. 2.5.
General proprioception
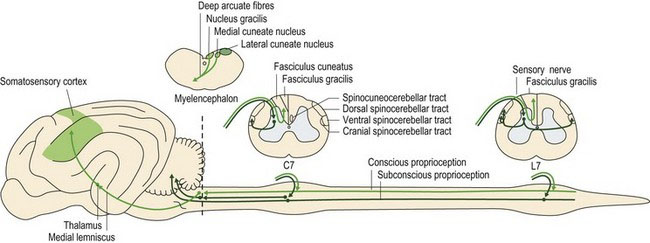
Proprioceptive information from peripheral receptors travels via spinal nerves, the dorsal roots and spinal cord (dorsal and lateral funiculi) to the brain (Fig. 6.1). Proprioception of the head, its muscles and joints, uses mainly cranial nerves VIII and V to reach the appropriate brainstem nuclei. The information that terminates in the somatosensory cortex of contralateral cerebrum is used in conscious proprioception while that terminating in the ipsilateral cerebellum is used in subconscious proprioception. Conscious proprioception is the conscious awareness of body position and movement of body parts. It enables the cerebral cortex to plan and refine voluntary, learned movements. Subconscious proprioception is based around stretch and tension of muscles, tendons and ligaments, at rest and during movement, and spatial orientation of the body. The cerebellum needs this information to coordinate posture and locomotion. The vestibular system provides proprioceptive information (conscious and subconscious) about head position and movement. This input is fundamental for setting the balance and posture of the whole animal.
Conscious proprioception
Key points
 Conscious proprioception is the information about the position of the head, body and limbs that is received in the contralateral somatosensory cortex of the cerebrum and is used particularly for executing voluntary, skilled movement.
Conscious proprioception is the information about the position of the head, body and limbs that is received in the contralateral somatosensory cortex of the cerebrum and is used particularly for executing voluntary, skilled movement.
 Information from the body and limbs is transmitted via the spinal nerves and spinal cord.
Information from the body and limbs is transmitted via the spinal nerves and spinal cord.


Non-painful, conscious sensations can broadly be divided into two categories that share functional and anatomical features. One group comprises touch and pressure, and the other is composed of joint and muscle/tendon proprioception. Both are principally transmitted to the cerebral cortex via the dorsal column–medial lemniscal system involving primarily the dorsal funiculus of the spinal cord with some transmission in the lateral funiculus (see Fig. 4.5). The dorsal funiculus is also known as the dorsal column and it continues as the medial lemniscus in the brainstem.
Axons conveying this information enter the dorsal funiculus and join the fasciculus cuneatus (from the body cranial to T8) or fasciculus gracilis (caudal to T8). Fibres in the fasciculus cuneatus convey proprioceptive and discriminative touch. They synapse in the medial cuneate nucleus located in the caudal medulla oblongata (see Figs. A26–30). Only about one-quarter of the fasciculus gracilis fibres synapse in the nucleus gracilis in the medulla oblongata. Thus the dorsal funiculus is relatively unimportant in transmitting proprioception from the pelvic limbs in quadrupeds; it is important in humans. The majority of fibres entering the fasciculus gracilis leave it in the cranial lumbar region to synapse in the grey matter on interneurons, or relay neurons of other pathways, or even LMNs. Fibres arising from synapses at the base of the dorsal horn (e.g. the nucleus thoracicus (see Table 4.2) may continue cranially in the dorsal spinocerebellar tract (with collateral fibres travelling in the adjacent spinomedullary tract) to synapse near the nucleus gracilis (in nucleus Z) (Figs. A24, A27, A28). Post-synaptic fibres from these proprioceptive relay nuclei (gracile, medial cuneate and Z) decussate in the deep arcuate fibres of the medulla oblongata and continue rostrally in the medial lemniscus to the thalamus where they synapse. They are projected to the somatosensory cortex of the cerebrum via the internal capsule.
The role of the lateral funiculus in conveying pelvic limb proprioception to the cerebrum may be one explanation why cervical spinal cord compression can produce particularly marked pelvic limb ataxia. This is illustrated in Fig. 6.3 on ‘Wobbler’ horses and dogs (see accompanying text).
The dorsal column–medial lemniscal system is highly organised topographically and is quite specific with respect to the transmission of sensory and discriminatory information. In other words, there is little convergence (input) of other pathways onto the relay nuclei; this enables the animal to localise precisely a stimulus from the skin, e.g. the horse biting at a fly on its body. Similarly, the somatosensory cortex is somatopically arranged and the size of each area reflects the density of innervation (see Fig. 4.14).
There are two kinds of proprioceptive input from the head – that associated with head position and movement (see Chapter 8) and proprioception due to receptors associated with muscles of mastication, facial expression and eye movement. Muscle proprioception is also conveyed by a three-neuron system via all three branches of CN V (trigeminal nerve). The trigeminal ganglion at the base of the neurocranium is the equivalent of a spinal ganglion, containing somatosensory neuronal cell bodies. However, uniquely, the cell bodies of the primary afferent neurons concerned with muscle proprioception are found in the CNS, in the mesencephalic nucleus of V. The second stage fibres decussate, travel rostrally in the trigeminal lemniscus to the thalamus and on to the somatosensory cortex. The proprioceptive fibres in the mesencephalic nucleus of CN V also connect to other cranial nerve motor nuclei for reflex function. Sensory fibres from tongue musculature (intrinsic and extrinsic) have their cell bodies located in the trigeminal, C1 and distal vagal ganglia.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


