Applied Digestive Anatomy and Feeding Behavior
Applied Anatomy and Function
Forestomach Anatomy
A comprehensive description of forestomach anatomy, secretory functions, and motility is found in Chapter 40, Disorders of the Digestive System.
Fermentation and Forestomach Microbes
Very little difference exists in the anaerobic microbial population between camelids and ruminants.1 However, Morvan et al. reported that llamas have greater acetogenic bacterial populations and also that camels have a total bacterial count lower than that of ruminants.2 Similarities between SACs and ruminants with regard to the forestomach fermentation process can be attributed to microorganism populations present in the fermentation chamber; these microorganisms determine the fermentation patterns and are primarily the result of diet composition and forestomach retention time.3
No major differences between SACs and ruminants exist in the volatile fatty acids (VFAs) produced during food fermentation. Concentrations of acetate, propionate, and butyrate of 68%, 19.7%, and 12%, respectively, have been reported for llamas fed at maintenance levels. The rapid rate of VFA absorption observed in SACs, which may be two or three times that of the rumen in ruminants, is an important factor that stabilizes forestomach pH favoring continued microbial fermentation and thus VFA production and availability.4
The forestomach fermentation process is further enhanced in SACs by physiologic adaptations related to urea recycling and liquid passage rate. SACs have a tremendous capacity to survive by consuming feed with low nitrogen as a result of higher nitrogen recycling efficiency through the forestomach wall, thus maintaining higher ammonia-nitrogen (NH3-N) content in the stomach compartments C1-C2 for microbial synthesis.5 Cellulolytic bacteria have an exclusive requirement for ammonia as their nitrogen source supporting microbial protein production.6 Another important factor in the digestive capacity of SACs is the higher forestomach liquid dilution rate in SAC compared with ruminants (see next section). A greater liquid dilution rate is indicative of greater bacterial production and arrival in the small intestine of SACs. The increase in the velocity of the liquid phase causes a greater efficiency of microbial growth in the C1-C2, ensuring that a minimum amount of energy is needed to maintain the microbial population.7–9
Consumption, Digestibility, and Feeding Behavior
Food Retention Time
Comparative studies between SACs and sheep on food retention time in the digestive tract show that SACs maintain the food for a longer time (Table 8-1).9,10 Also, Clemens and Stevens and Sponheimer et al. found that SACs retain large particles for a longer period compared with other species of mammals such as cattle, horses, goats, and rabbits.11,12 The longer retention time in SACs has two important consequences: (1) the improved utilization of cell wall constituents, and (2) the restriction of consumption by limiting forestomach capacity.13

With regard to the liquid phase rate of passage in C1-C2, SACs have a faster rate of passage compared with sheep: 10.4% per hour compared with 7.7% per hour (Table 8-2).9 Also, Clemens and Stevens found that llamas have a faster dilution rate of fluids and smaller particles compared with cattle and horses.11 Maloy observed a faster liquid phase flow in the stomach of camels than in that of Zebu steers.14
TABLE 8-2
Forestomach Volume and Estimate of Liquid Passage Rate in Llama and Sheep
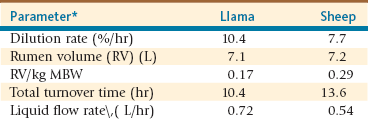
*kg, Kilogram; L, liter; L/hr, liter per hour; MBW, metabolic body weight = BW kg0.75; %/hr, percent per hour.
Mechanisms accounting for differences in the retention times of liquid and solid contents in the forestomach of ruminants are not clearly understood. The probability of feed particles leaving the forestomach is mainly determined by density and size, although particle density is more important than size. Larger particles have low density and are therefore retained for a longer period in C1-C2.13 Prolonged retention of larger particles facilitates continued microbial cellulose degradation.
The rapid liquid passage rate in SACs promotes microbial production by forcing microbial populations to maintain active growth (i.e., log growth). The active growth phase of microbial populations results in greater efficiency of microbial growth in the C1-C2, ensuring that only a minimal amount of energy is needed to maintain the microbial population.7,15
Consumption
Most available information on consumption in SACs comes from comparative studies in stabled conditions. Dry matter consumption observed in alpacas and llamas is 1.8% and 2.0% of live weight, respectively (Table 8-3). In alpacas and llamas, the consumption of organic matter gram per kilogram of metabolic body weight (g/kg BW0.75) ranges from 40 to 56. In general, the daily feed consumption by SACs is less than that of sheep. San Martín found that under grazing conditions, llamas and alpacas have the same level of consumption, being approximately 36% and 26% lower than that seen in sheep under cultivated and native pastures, respectively.9 The lower consumption rate observed in SACs in comparison with ruminants is a result of longer particle retention time in the digestive tract of SACs.16,17
TABLE 8-3
Comparative Daily Intake among Alpaca, Llama, and Sheep under Housed and Grazing Conditions
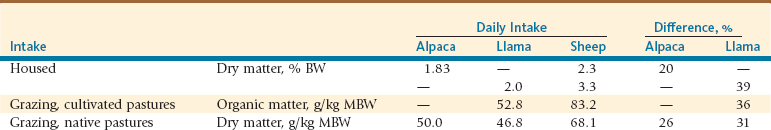
g/kg, Gram per kilogram; MBW, metabolic body weight = BW kg0.75
In the Altiplano grasslands, the SAC feed consumption during the dry season is similar to or greater than consumption during the rainy season despite the lower nutritional quality of native pasture in the dry season compared with the rainy season. This level of consumption in the dry season is in response to the increase in gastric capacity because of the low nutritional quality of the diet. This is observed in other species as well.18–20
In animal production, low intake is often related to low rate of production because a large proportion of the energy from food is diverted to meet maintenance requirements, which results in poor feed conversion. In contrast, the low consumption observed in SACs managed in their harsh native environment with low availability of poor-quality forage during most of the year is an important adaptation, which, coupled with greater digestive capacity, provides competitive advantage.
Digestibility
Several comparative studies addressing in vivo digestibility have been conducted in SACs and ruminants. San Martín et al. reviewed several in vivo digestibility trials in alpacas and sheep, in which animals were separated into groups on the basis of the dietary crude protein content being less than 7.5% or greater than 10.5%.21 For diets containing less than 7.5% crude protein, digestibility was found to be higher and more favorable to alpacas, whereas no difference between species was observed in diets with levels higher than 10.5% crude protein. Additional studies on comparative digestibility have confirmed these findings (Table 8-4). Thus, SACs show higher digestibility compared with ruminants consuming low- to medium-quality diets, whereas comparable digestibility was observed between the two species for diets of high quality or low fiber content.9,12,17,22
TABLE 8-4
Differences in Organic Matter Digestion between Llamas and Sheep in Relation to Diet Quality
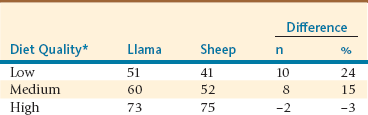
*Low = 7% crude protein (CP) and 2.2 megacalorie (Mcal) digestible energy (DE) per kilogram of dry matter.
San Martín et al. and Genin and Tichit, using a dacron bag technique to estimate in situ digestibility between SACs and ruminants, observed greater digestibility in SACs compared with ruminants, even with equal retention time in both species.23,24 These results indicate that other factors besides retention time are implicated in the digestive superiority of SACs. These include increased frequency of forestomach contractions, ruminant cycles, high ratio of salivary flow to stomach volume, and presence of forestomach glandular saccules. Additionally, greater digestibility of low-quality food is facilitated by the ability of SACs to maintain a higher concentration of NH3-N in C1-C2 compared with ruminants. This would provide more available nitrogen for microbial synthesis, improving fiber digestion.5,25
Feed Selectivity
The lips of the SACs are relatively thin. The upper lip is divided by a median groove (cleft lip) and is smaller than the lower lip. The SAC lips are mobile, which allows for high selective ability. SACs do not use their tongue to manipulate food, and it rarely protrudes from their mouth, making them unlikely to lick salt licks.4
Premolars and molars play an important role in efficiency of cutting and grinding of food. During mastication, vertical and horizontal mandibular movements allow for efficient grinding of food leading to a reduction of particle size.26 Florez and Gutierrez showed that alpacas and llamas are more efficient at reducing particle size compared with sheep.27 In llamas and alpacas, efficiency in particle size reduction increases when forage length increases, whereas the opposite was observed in sheep.
Diet Composition
Early studies on diet selectivity in alpacas were completed on the Peruvian rangelands by direct observation. Alpacas eat mostly tall grasses in the rainy season and short grasses during the dry season. Festuca dolichophylla, Distichia muscoid, Trifolium amabile, and Bromus unioloides were identified as the preferred species.28 Bryant and Farfan, studied alpacas grazing in the Festuca-Calamagrostis grasslands using microhistologic analysis of fecal material and observed that alpacas were primarily grazers rather than forb eaters during the dry season and early wet period.29
Some studies on diet selectivity used alpacas with esophageal fistulas. In the Festuca-Calamagrostis grasslands, the selected plant species during the rainy season were Hipochoeria estinophala (18%), Trifolium amabile (16%), Nothoscardium sp. (16%), and Eleocharis albibracteata (15%) and in the dry season Festuca dolichophylla (56%) and Calamagrostis vicunarum (28%).30 In the Festuca-Stipa grasslands, it was reported that selection of short grasses increased, whereas tall grass selection declined in the last months of the dry season. The dominant species in the diet were Festuca dolichophylla (21%), Muhlenbergia peruviana (20%), Festuca rigida (17%), and Stipa brachiphylla (15%).31 Alpacas showed a high degree of preference for wetland when restricted to high altitude. This wetland is characterized by sedges (Carex crespitosa) and green vegetation; alpaca diets consists mainly (78%) of ciperaceas (sedges) and juncaceas (rushes). In contrast, in grassland dominated by Festuca dolichophylla and Muhlenbergia fastigiata, diets were mainly (60%) composed of grasses.32
In the highlands of Peru, about 70% of the livestock are handled by small farmers and communities. Also, llamas and alpacas are raised together with sheep. Despite this fact, few studies have focused on comparisons of diet composition between these species to understand the differences in the utilization of feed resources within the pastoral system. Pfister et al. reported that in their study, llamas spend more time at each feeding station than do alpacas or sheep and that SACs have greater total grazing times compared with sheep.33 However, Sharp et al. reported that alpacas in New Zealand show more grazing activity compared with sheep during daylight and less activity at night, but significantly more ruminating activity during the night.34
San Martín reported a comparative botanical composition of diets consumed by llamas, alpacas, and sheep grazing under three different cultivated pastures (Festuca rubra, Lolium perenne, and Trifolium repens) and two native grasslands dominated by Festuca dolichophylla and Festuca rigida.9 In the cultivated pasture, sheep consumed about 2.6 times more legumes compared with SACs. The legumes were selected less by SACs, which may explain why these animals do not experience bloat as readily as sheep do. Tall grasses were selected more by llamas when grazing native grasslands. Also, llamas had a lower selection of leaves compared with alpacas and sheep.
These observations suggest that llamas are adapted to arid environments, which is further supported by their current distribution. Approximately 70% of the world’s llama population is located in the Bolivian highlands, where rainfall fluctuates between 250 to 450 millimeters (mm) (9.8 to 17.7 inches). In contrast, the Peruvian Altiplano contains only 25% of the world llama population, and rainfall in that region fluctuates between 500 and 900 mm (19.7 and 35.4 inches). Within the Peruvian Altiplano grasslands, the llama population is located mainly in arid areas (dry puna).35 Additionally, it was observed that llamas are susceptible to foot rot problems when grazing on wet soils, a problem that rarely occurs in alpacas.
Shimada and Shimada suggested, on the basis of ethnographic, archeozoologic, physiologic, and ethnohistoric data, that llamas were bred and maintained on the north coast of Peru as early as 600 BC.36 They indicated that llamas are physiologically adapted to arid environments and that current absence of these animals in the Peruvian coast has resulted from competition with animals introduced by the Spaniards. It is interesting to note that selective characteristics observed by San Martín resemble those seen in the Old World camels.9 The Old World camels prefer sparse vegetation and dry grasses that are usually avoided by other animals.37 Also, Genin et al. suggested that llamas prefer dry, tall, and fibrous forages more than do other ruminants.38
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


