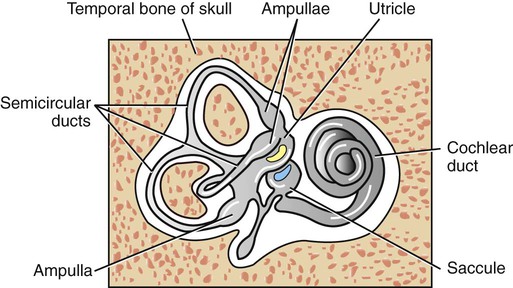
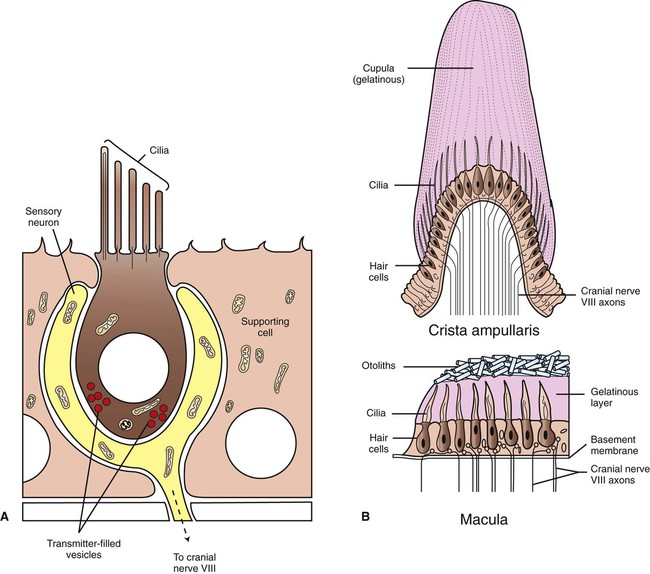
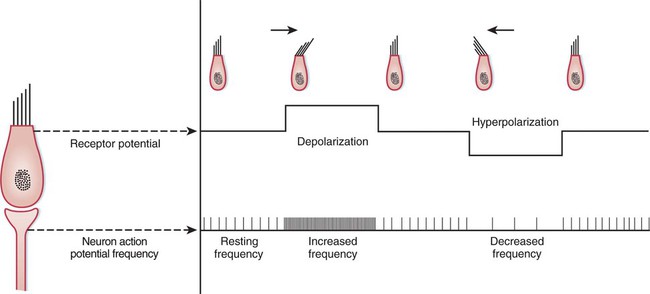
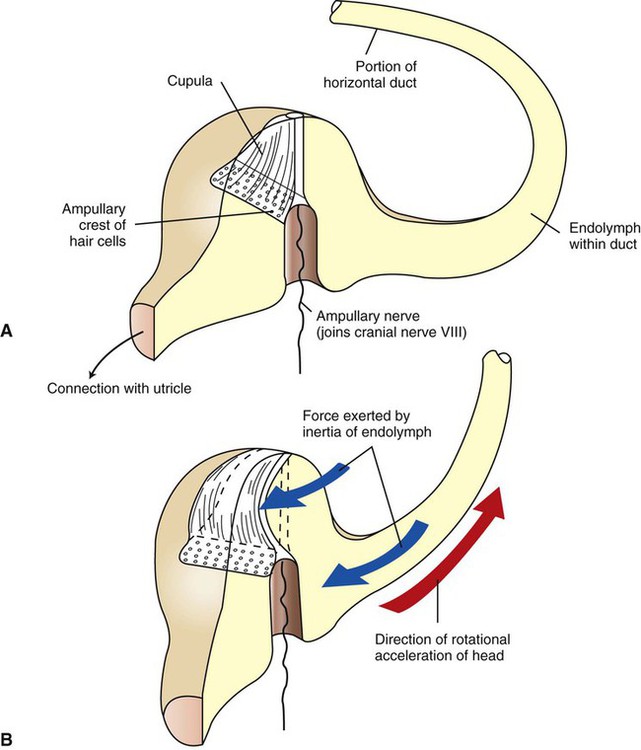
1. The vestibular system is a bilateral receptor system located in the inner ear. 2. Specialized regions of the vestibular system contain receptors. 3. The semicircular ducts detect rotary acceleration and deceleration of the head. 4. The utricle and saccule detect linear acceleration and deceleration and static tilt of the head. 5. The vestibular system provides sensory information for reflexes involving spinal motor neurons, the cerebellum, and extraocular muscles of the eye. 6. Vestibular reflexes coordinate eye and head movements to maximize visual acuity. To coordinate posture and locomotion, the brain needs to know not only what movement it intends to command, but also the orientation of the body and what movement the body is actually performing. Chapter 8 describes the muscle spindle, an important source of information for the brain about body position and movement. Another important source of information is the vestibular system. This bilateral receptor system is located in the inner ear and informs the brain about the position and motion of the head. The inner ear, or labyrinth, is made up of two parts: the bony labyrinth and the membranous labyrinth. The bony labyrinth is a system of caverns and tunnels through the petrous temporal bone of the skull. The bony labyrinth houses the receptor organs of the vestibular system as well as the receptor organ for hearing, the cochlea (Figure 11-1) (see Chapter 17). These receptor organs are part of the membranous labyrinth, which consists of thin membranes of epithelium and lies within the bony labyrinth. This epithelial membrane is specialized at some locations to become the sensory receptor cells that form the vestibular and auditory receptor organs. The membranous labyrinth is filled with a fluid called endolymph and is separated from the bony labyrinth by a fluid called perilymph. The vestibular portion of the membranous labyrinth consists of two major sets of structures: (1) three semicircular ducts, located at approximately right angles to each other, and (2) a pair of saclike structures called the utricle and saccule, sometimes called the otolith organs. As discussed later, each major set of structures is respectively involved in transducing a different major class of vestibular stimulus. Each vestibular structure of the membranous labyrinth has a region of epithelial lining that has become specialized into a set of secondary receptor cells (see Chapter 7) called hair cells (Figure 11-2). These hair cells form the basis of a sensory receptor organ within each vestibular structure. Each hair cell has several cilia at its apex that are arranged in order according to size. At its base the hair cell synapses with a sensory neuron that carries action potentials to the brainstem. The cell bodies of these sensory neurons are located in Scarpa’s ganglia, and their axons collectively form the vestibular portion of the vestibulocochlear nerve (cranial nerve VIII). The cilia from all the hair cells within any one vestibular structure project into a gelatinous mass; displacement of this gelatinous mass in a given direction causes all the hair cell cilia to bend in that direction. At rest, when the cilia are not deflected, the sensory neurons that synapse with the vestibular hair cells transmit action potentials spontaneously at about 100 per second (Figure 11-3). When the hair cell cilia are bent in a direction toward the largest cilium, the hair cells depolarize, the release of transmitter from the hair cells onto the sensory neurons increases, and the action potential frequency of the neurons increases. When the cilia are bent in the opposite direction, toward the smaller cilia, hair cell membranes hyperpolarize, transmitter release decreases, and the action potential frequency of sensory neurons decreases. Therefore, displacement of the hair cell cilia in either of these directions can be detected by the brain as an increase or a decrease from the resting action potential frequency. Deflections in other directions are much less effective. How the brain uses this information to detect the direction of head movement is described later. Three membranous semicircular ducts are located within corresponding semicircular canals of each bony labyrinth (Figure 11-4). They are positioned at approximately right angles to each other, and both ends of each fluid-filled duct terminate in the utricle. Each semicircular duct has an enlargement at one end, called the ampulla, near its junction with the utricle. The ampulla contains a hair cell receptor organ called the crista ampullaris (see Figure 11-2). This is a ridge of hair cells that, at their base, synapse on sensory neurons projecting to the central nervous system (CNS) and whose cilia are embedded in an overlying gelatinous mass. This mass, called the cupula, attaches to the roof of the ampulla (Figure 11-5). All hair cells of a given crista ampullaris are oriented in the same direction with respect to their cilia. Together, the hair cell ridge and overlying cupula span the diameter of the ampulla.
The Vestibular System
The Vestibular System Is a Bilateral Receptor System Located In the Inner Ear
Specialized Regions of the Vestibular System Contain Receptors
The Semicircular Ducts Detect Rotary Acceleration and Deceleration of the Head
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


