4 Water
The desire for water is a basic motivating instinct for animals which is only exceeded by their need for air and the avoidance of severe pain. Water is a universal solvent therefore, “water is involved either directly or indirectly in virtually every physiologic process essential to life” (Raisbeck et al 2007). Water is critical to all biochemical reactions and thermoregulation. Water reserves in horses are more labile than energy reserves (body fat), which allows horses to tolerate feed deprivation for a longer period than a lack of water. Dehydration greater than 15% percent can be fatal for a horse (Carlson 1979). This is a fluid loss of about 95 liters for a 500 kg horse. Depending on environmental conditions and activity, fluid loss to this extent, as a result of obligatory fluid losses (Fig. 4.1), would be likely to take less than 7 days. Death due to starvation occurs after a body mass loss of about 50% (Stull, unpublished information). Depending on many factors (age, body condition, climate, activity), a weight loss of this magnitude for an average horse would likely take more than 90 days.
Body fluid compartments
Total body water accounts for 61–72% of the body weight (BW) of mature horses (Table 4-1: Julian et al 1956, Deavers et al 1973, Carlson et al 1979, Judson & Mooney 1983, Carlson 1987, Sneddon et al 1993, Forro et al 2000, Fielding et al 2004, 2007, 2008, Lindinger et al 2004, Waller & Lindinger 2005, Forro & Lindinger 2006) and has been estimated at 66–84% BW of foals varying with foal age or dilution method used (Oftedal et al 1983, Doreau et al 1986, Geerken et al 1988).
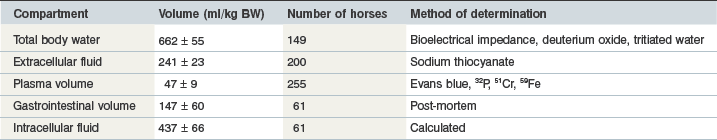
Total body water (TBW) is distributed within cells (intracellular) or outside of cells (extracellular). The intracellular fluid compartment (ICF) is estimated at 38–53% of body weight (BW) (Table 4-1). The extracellular fluid compartment (ECF) comprises fluid in blood, interstitial fluid (ISF), bone, connective tissue, and transcellular fluid. The ECF compartment has been estimated at 22–26% BW (Table 4-1; Andrews et al 1997, Fielding et al 2003, 2004, 2007, 2008, Waller et al 2008). Transcellular fluids are contained within epithelial-lined compartments such as the gut and urinary bladder. Depending on diet, the fluid sequestered by the horse’s gastrointestinal content can account from 9 to 21% of equid weight (Robb et al 1972, Coenen & Meyer 1987, Gee et al 2003, Sneddon et al 2006) and has been speculated to be a fluid reservoir for the horse during physical activity and other brief periods when water is inaccessible (Sneddon & Argenzio 1998). The fluid holding capacity of the diet influences the water content in the equine gastrointestinal tract. Hay-fed horses had a total intestinal fluid capacity of 188 ml/kg BW which was 84% greater than horses fed a complete feed (102 ml/kg BW) (Coenen & Meyer 1987). Moreover, 77% of gut water in hay-fed horses was in the large bowel compared to 65% in horses fed complete feed. The large reservoir of gut fluid may explain why horses drink intermittently rather than sip continuously.
Total body water decreases linearly with age in horses and foals (Agrabriel et al 1984, Doreau et al 1986). The ECF of newborn foals is about 40% of BW but declines to 25% of body weight in adult horses (Table 4-1; Spensley et al 1987). Likewise, plasma volume in newborns is higher (9.6% BW) (Table 4-1) than in mature horses (4.7% BW) (Spensley et al 1987). These differences underscore why the approaches for fluid therapy in the foal must differ to that used for the mature horse.
Water balance
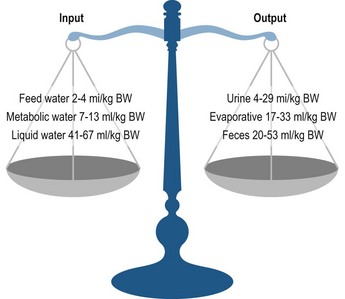
Water balance describes the homeostasis between water input and water output (Fig. 4.1). Water input occurs through drinking, water contained in feed, and water derived by metabolic processes. All horses lose fluid via four routes: fecal, urinary, respiratory, and cutaneous (sweat) losses. The latter two routes are called evaporative heat loss. Lactating mares have a fifth route of body fluid loss via milk secretion. The water balance equation for horses can be given as:
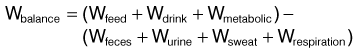
Wmilk is added to the equation for lactating mares. Two criteria are difficult to measure: metabolic water and water in respiration. Measuring cutaneous losses is difficult but not impossible. Although the water balance diagram (Fig. 4.1) implies a static relationship, the association between water intake and water output is dynamic.
Fluid output or loss
Fecal fluid loss
Feces are the main route of fluid loss in the mature, idle horse fed hay (Table 4-2) although diet, environment, physical activity and husbandry have a marked influence on absolute amounts of fluid lost by this route. Diet is one of the main determinants of intestinal, and therefore, fecal water content. Horses fed hay or silage (72–85% moisture) have wetter feces than horses fed hay diets with added grain (66% moisture) (Fonnesbeck 1968, Cymbaluk 1990a, Meyer 1995, Warren et al 1999, Zeyner et al 2004, Muhonen 2008). Absolute fecal fluid losses range from 16 to 38 mL/kg BW (Tasker, 1967c) (also see Table 4-1). Based on retrospective analysis of fecal moisture content of horses fed various hays (timothy, alfalfa-grass, oats, hay) plus limited grain mixes (less than 15%), fecal fluid output averaged 31.0 ± 4.9 ml/kg BW (range 19.6–52.7 ml/kg BW) (Cymbaluk unpublished data).
Table 4-2 Diet Effect on Water Intake, Fecal and Urinary Fluid Output of Idle Horses Kept in a Thermoneutral Environment (10°C, RH <75%)
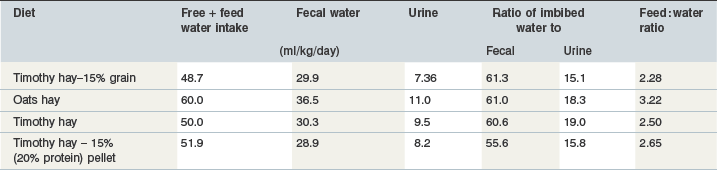
Source: N.F. Cymbaluk, unpublished data.
Fecal moisture content and total fecal water output were positively correlated to total DM intake and crude fiber content over a range of fecal moistures of 66 to 79% (Fonnesbeck 1968, Cymbaluk 1989). Water content of ingesta of horses unadapted to grain then acutely fed two high grain meals (4.55 kg/day) was approximately 5% lower than for horses fed an all hay diet free choice yet fecal water content did not differ significantly (Lopes 2002). Drier manure (less than 70%) was observed in horses fed high grain (greater than 50%), high glucose (15%), and high molasses beet pulp (26%) diets (Cymbaluk 1989, Olsman et al 2004). However, horses fed a high (12%) soluble fiber diet derived from beet pulp and alfalfa hay had a fecal moisture content (81.7%) 8% wetter than those fed a low (7%) soluble fiber diet of oats-orchardgrass–timothy hay as a result of a higher water : feed intake and adsorption of water to hydrophilic fiber (Warren et al 1999). Moreover, horses fed an all-hay diet maintained lower plasma protein concentration post-exercise than horses fed a limited hay–concentrate diet, which was inferred to represent conservation of plasma volume through greater movement of fluid from the gut (Danielsen et al 1995). For this reason, a higher dietary soluble fiber content has been speculated to have possible benefit during exercise when there is an increased water need. Although the pretreatment plasma volume of horses fed a high soluble fiber diet was higher, consuming a high or low soluble fiber diet had no effect on the plasma volume of horses after frusemide administration (Warren et al 1999). Similarly, no differences were observed in hydration status following a 60 km exercise test in horses fed a chopped grass hay compared to a grass hay : soluble fiber chop (Spooner et al 2009).
Intestinal disease that impairs water reabsorption by the gut increases fecal water loss. Horses with chronic diarrhea had a fecal output nearly fourfold greater (50.7 kg/day) than normal horses (12.04 kg/day) (Merritt 1975). The large fecal fluid loss by horses with chronic diarrhea was compensated for by a doubling of water intake (42 vs 24 liters/day). Likewise, horses with castor oil-induced diarrhea had fecal moistures of 90% or higher and lost twice the volume of fecal water (5.15 ml/kg BW/h) of normal horses (2.15 ml/kg BW/h) (Ecke et al 1998). For a 500-kg horse, these values equate to a daily fecal fluid loss of 61.8 liters/day during diarrhea compared to 25.8 liters/day in the normal horse. It is evident why horses with diarrhea become dehydrated if fluids are not replenished.
Urinary fluid loss
Body fluid homeostasis is controlled by the kidney. Although specific gravity (SG) alone has been used to classify urine concentration (Savage 2008), use of urine osmolality and urine SG provide a more accurate definition (Wilson 2007). Hyposthenuria describes urine with a SG less than 1.008 and an osmolality below 269 mOsm/kg (Wilson 2007). Isosthenuric urine is similar to plasma in concentration with an SG of 1.008–1.012 and osmolality between 260–300 mOsm/kg. Hypersthenuric urine has a SG greater than 1.012 with an osmolality greater than 300 mOsm/kg. Normal adults have hypersthenuric urine. Normal foals have hyposthenuric urine (Wilson 2007).
The obligatory loss of urine is the fluid required to eliminate various solutes via the kidney irrespective of fluid intake. Horses have an obligatory urine loss of about 5–6.3 ml/kg BW per day (Tasker 1967a) whereas “normal” urine volumes range from 6 to 29 ml/kg BW (Fonnesbeck 1968, Rumbaugh et al 1982, Freeman et al 1999). The average daily urine output for idle horses (n = 340) housed in a thermoneutral environment and fed a wide range of feeds was 8 ml/kg BW (range 4–18 ml/kg BW) (Cymbaluk, unpublished data). Variance in normal urine volume arises from differences in diet (composition and nutrient content), water or fluid availability, metabolic responses to ambient temperature, exercise load or gastrointestinal health (Tasker 1967b, c, Cymbaluk 1989, Knottenbelt 2000, Connysson et al 2006). Chronically diarrheic horses (4.15 liters/day) reduced urine output by 23% compared to normal horses (5.36 liters/day) (Merritt 1975). During and after heavy exercise, horses reduce urine output to conserve body fluid. Exercised Thoroughbred horses produced 1.4 liters less urine per day than idle horses perhaps to compensate for fluid losses in sweat (Pagan et al 1998). Yet, the impact of exercise on urine volume is unpredictable because fluid conservation is tied to sweat loss which depends on exercise intensity and the environmental conditions in which the exercise is performed.
Urine output in the horse varies quickly with changes in physiological and biochemical stimuli. Changes to the renal solute load created by protein, sodium, or potassium above the horse’s requirements typically elevates urine volume. Urine flow rates quadrupled in donkeys fed alfalfa hay compared to a similar dry matter intake provided by a wheat straw diet because of an 8-fold higher nitrogen intake on the alfalfa diet (Izraely et al 1989). Because potassium homeostasis is regulated by the kidney, horses fed a high potassium diet (5.4 mmol K/kg BW) had a 26 to 30% higher urine output than horses fed a diet with 4.1 mmol K/kg BW (Jansson 1999, Jansson & Dahlborn 1999, Jansson et al 1999). Urinary output may be expected to increase when diets containing high-potassium feeds such as molasses (6.1% potassium) are fed to horses (NRC 2007). Although sodium is effectively conserved by the kidney, regulation of equine sodium homeostasis may be gastrointestinal rather than renal (Jansson & Dahlborn 1999). This is supported by data showing that ponies had similar urine volume when fed high (1–%) dietary salt (Schryver et al 1987).
Total evaporative fluid losses
Heat loss in horses occurs by water vaporization through respiration and/or sweating. At body temperature, 2.5 kJ or 578 calories of heat are removed during evaporation of 1 ml of water. Evaporative heat and, therefore, fluid loss, occurs by passive diffusion from skin and lungs whereas sweating is an active process, initiated by elevated body core temperatures, involving fluid secretion by sweat glands. Sweating occurs at thermoneutrality but increases greatly when core body temperatures increase in response to exercise or elevated ambient temperature (Lindinger 2008). The distribution of total evaporative heat loss in exercised horses is about 23% respiratory and 70% or more through sweating (Kingston et al 1997). Daily evaporative losses of idle Standardbred horses fed alfalfa hay and kept in thermoneutral (5–20°C) conditions were calculated at 10 liters/day with an average of 23 ml/kg BW (Groenendyk et al 1988). Standardbred horses kept in a climatic chamber had evaporative heat losses of 48 W/m2 or about 20 ml/kg BW at thermoneutral temperatures (Morgan et al 1997).
Respiratory losses
Respiratory heat (fluid) loss by the idle horse varies with ambient temperature and humidity. In the exercised horse, fluid losses increase with length and duration of exercise. Respiratory fluid loss accounted for 19 to 30% of the total evaporative heat loss by exercised horses (Hodgson et al 1993, Kingston et al 1997). Thoroughbred horses exercised at 40, 65, and 90% of VO2max for durations of 38, 15, 9 minutes, respectively, had estimated respiratory fluid losses of 2.1, 0.8 and 0.8 liters and concurrent sweat losses of 9, 9.2, and 6.9 liters, respectively (Hodgson et al 1993). Respiratory heat loss by Thoroughbred horses undergoing long duration exercise (202 min) at 40% VO2max was estimated at 23% of evaporative heat loss (7.4 liters) with concomitant sweat losses of 22.7 liters (Kingston et al 1997). Heat acclimation occurs through increased sweating and respiration as discussed in more detail below (Geor et al 2000, McCutcheon & Geor 1999).
Sweat losses
Passive cutaneous evaporative loss by horses (average weight 429 kg) housed at temperatures between 5 to 20°C in a climatic chamber were estimated at about 8.5 liters per day (see above) (Morgan et al 1997) but in exercised horses, total fluid losses increase through sweat in proportion to exercise duration and intensity, environmental conditions, and climatic acclimation of the horse (McCutcheon & Geor 1998). Rates of fluid loss by horses doing low (40% VO2max), moderate (65% VO2max) and high (90% VO2max) intensity treadmill work in an air-conditioned laboratory (21–23.5°C) were 0.61, 1.32 and 1.73 ml/kg BW/min (Hodgson et al 1993) Total body water losses attributed to sweating by cross-country event-horses ranged from 4.8 to 20.4 liters (2–6.1% of BW). (Ecker & Lindinger 1995). Body mass losses of 33.8 kg were obtained for 460-kg horses performing long-distance (45 km), low-intensity (40% VO2max), activity on a treadmill (Kingston et al 1997).
Training, conditioning and temperature acclimation increase tolerance to high ambient temperatures. Training alone increases sweating rate of horses which is intensified as conditions of high humidity and high ambient temperatures are imposed (McCutcheon & Geor 1999). High ambient temperatures alone (>33–35°C) increased evaporative heat (fluid loss) by four- to five-fold in idle horses (Morgan et al 1997) and by 45–60% in exercising Standardbred and Thoroughbred horses relative to fluid loss at 20°C (Jansson 1999, McCutcheon et al 1995).
Sweating, in response to hot weather, requires time to become fully effective. Partial acclimation by horses to hot weather occurs within 2 weeks whether hot temperature exposure is intermittent or continuous but 3 weeks of exposure is required for full acclimation (Geor et al 2000, McCutcheon & Geor 1999). Acclimation to hot ambient temperatures does not confer resistance to dehydration.
Lactation losses
Milk output varies widely among individual mares, breeds, and over the lactation cycle (Doreau & Boulot 1989). Typically, primiparous mares produce less milk than multiparous mares irrespective of mare breed (Doreau et al 1991, Pool-Anderson et al 1994). Quarter horse mares produced 18–21 ml milk/kg BW daily (Gibbs et al 1982, Pool-Anderson et al 1994), Thoroughbred and Standardbred mares yielded 29 to 34 ml/kg BW per day (Oftedal et al 1983), primiparous and multiparous light and draft mares produced 24 to 27.5 ml milk/kg BW per day (Doreau et al 1991), Australian stock horses produced 37 ml/kg BW per day (Martin et al 1992), and Lusitano mares produced 23–30 ml/kg BW per day (Santos & Silvestre 2008). To offset lactational losses only, mares must increase water intake by 30 to 60% above maintenance.
Key Points –
Water losses
• Feces are often the main route of fluid loss in idle horses but this depends on diet. Average daily fecal fluid output is about 31 ml/kg BW with a range between 20 to 53 ml/kg BW.
• Daily evaporative losses for horses average about 20 ml/kg BW (range 17–33 ml/kg BW) at thermoneutral conditions. Sweat fluid losses increase markedly with exercise, with further increments when exercise is conducted in hot conditions.
• Daily milk losses by mares range from 18 to 37 ml/kg BW under thermoneutral conditions. Water intake can be expected to increase by 30 to 60% to offset lactational losses alone. The metabolic requirements of lactation result in the consumption of at least 36% more feed during lactation than during pregnancy, thus water intake will increase proportionately to the increase in feed intake. Hot weather will further elevate water intake requirements.
Water intake
Water provision can be external (direct) or internal (indirect). External fluid supply is provided by liquid (free) water and feed water. Imbibed water generally supplies more than 80% of the water requirements of hay-fed horses whereas grazing horses obtain significant amounts of water from pasturage. Metabolic water is generated by tissue oxidation of protein, fats, and carbohydrates from feed and is estimated at 11–13% of daily water intake or 6.8–13 ml/kg BW (Groendyk et al 1988, Van den Berg et al 1998) and has been estimated at about 2.7 liters/day in a 500 kg horse (Carlson 1979).
Feed water
All horse feeds contain water. Basal feeds used in the stable such as hay, grain and processed feeds typically contain 10 to 15% moisture and so supply little fluid (1 to 2 liters/day) to the horse’s water balance (Freeman et al 1999, Muhonen 2008). Haylage (32–45% moisture) and silage (55–64% moisture) can supply about 25% and 45% of the water needs of idle and worked horses resulting in a proportionate decrease in volume of liquid water consumed when these feeds are fed (Muhonen 2008). A vegetative perennial ryegrass pasture (80% moisture) in the amounts eaten by pregnant (68 kg) and barren mares (40 kg) provided 54.5 kg and 31 kg feed fluid, respectively (Marlow et al 1983), and nearly met the mares’ water needs. Pasture moisture content declines over a growing season from more than 80% in the spring to less than 10% by late fall (McMillen et al 1943, Meissner & Paulsmeier 1995). This explains why equids drink infrequently (1.4 to 3.4 bouts/day) on lush pasture (Scheibe et al 1998) but drink more often as pastures senesce and when dry feeds are provided in stables (Sweeting et al 1985).
Liquid water
The total volume of water consumed by an idle horse is determined by its body weight. Therefore, an idle 1000-kg Percheron horse will typically drink more water than a 600-kg Quarter horse (Fig. 4.2). Adult horses fed hay to meet maintenance energy needs drink about 50 ml/kg BW daily. Daily free water intake by normal, idle horses fed alfalfa-timothy or alfalfa hay ad libitum was 51 to 56 ml/kg BW (Tasker 1967b, Sweeting et al 1985, Groenendyk et al 1988), stabled grass-fed Namib and Boerperd horses drank 41 to 48 ml water/kg BW (Sneddon et al 1993), stabled, mature ponies fed grass or alfalfa hay drank 50 to 55 ml/kg BW (Cymbaluk 1989), pregnant mares fed a grass hay-grain diet drank 45 to 59 ml water/kg BW (Freeman et al 1999).
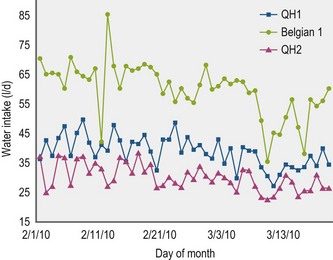
Horse breed may affect water intake but this factor has been inadequately tested. Thoroughbred horses and Highland ponies drank twofold more water per kg feed than donkeys or Shetland ponies (Cuddeford et al 1995). Draft-Thoroughbred cross mares (55.2 ± 11.5 ml/kg BW) fed the same diet drank 13% more water than Quarter horse mares (48.7 ± 7.75 ml/kg BW) (Cymbaluk, unpublished data) (Fig. 4.3) and donkeys drank 27% less water (51 ml/kg BW) than similarly housed and fed ponies (65 ml/kg BW) (Mueller & Houpt 1991).
Although daily maintenance water requirements are estimated at 50 ml/kg BW (range 41–67 ml/kg BW), individual horses of similar weight fed similar diets can drink very different amounts of water. Not only do individuals of similar breeds differ, an individual horse varies its water intake from day to day (Fig. 4.2) (Groenendyk et al 1988), likely because horses have a cyclic pattern of DM intake over time and may vary their intake of dietary ingredients (Jansson & Dahlborn 1999). Absolute water intake by pregnant mares varied 16 to 20% from the average (Freeman et al 1999) and by 21 to 25% in Standardbred horses (Nyman et al 2002). Daily water intake by individual Thoroughbred mares varied up to 75% (Smith et al 1996).
Diet effects on water intake
The total amount of feed eaten and its composition alters total water intake by the horse (Fonnesbeck 1968). Ponies drank 12 to 61% more water when fed all-hay diets than when fed mixed grain-hay diets (Cymbaluk 1989). Although Thoroughbred horses fed a hay-cube diet drank 26% more water than when fed a 40 : 60 grain : forage diet, water : feed ratios were similar (3.87 for all forage versus 3.66 for grain : forage; Pagan et al 1998). Thoroughbred horses fed diets with 50% grain drank significantly less (p = 0.02) water (17.4 liters) per day than those fed a 35% grain (22.4 liters water/day) and defecate less frequently (p = 0.06) (Freire et al 2009). Welsh ponies fed perennial ryegrass-timothy in a chopped, dehydrated form drank 23.8% more water than when the same forage was fed in the long form despite similarity in total DM intake (Hyslop 2003). Water : feed ratios for mature cross-bred ponies fed grass hay, alfalfa hay or high grain complete feed pellets were 3.2, 3.3 and 2.0, respectively (Cymbaluk 1989), and were 3.4, 3.3 and 2.6 for horses fed alfalfa-beet pulp, 77% orchardgrass-alfalfa-oats and forage-grain diets, respectively (Warren et al 1999). Ponies fed alfalfa and oat straw in ad libitum quantities had water : feed ratios of 4.4 and 3.8, respectively, but donkeys drank less water per kg DM than ponies when fed alfalfa (3.2 liters/kg) or oat straw (3.2 liters/kg) (Pearson et al 2001). The latter study also showed that water : feed ratios increased when horses were limit-fed at 70% of ad libitum feed intakes. The higher water intake by horses fed higher forage was attributed to a higher fiber intake of those diets. Adding grain to the diet reduces the total amount of feed and fiber eaten, which may explain the lower water intake by horses fed high grain diets.
Dietary composition
Doubling the salt intake in a compound feed from 50 to 100 mg/kg BW, increased water intake by 53% and increased urine output by 47% (Meyer et al 1987). Water intake in one study was linearly correlated to salt intake (y = 36.5 + 0.22x where y = daily water intake in ml/kg BW and x = daily sodium intake in mg/kg BW) over a narrow range of intakes (Jansson & Dahlborn 1999). Others (Schryver et al 1987) have found no difference in water intake by ponies fed 1, 3, and 5% dietary salt.
Based on a limited range of protein intakes, water intake (WI, ml/kg BW) correlated linearly to nitrogen balance (N balance, mg/kg BW): N balance = 4.8339 WI − 54.696 (r2 = 0.95) (Ragnarsson & Lindberg 2008). Similarly, Hyslop (2003) reported a linear relationship between water and protein intake for Welsh pony geldings of WI (liters/day) = 19.61 + 0.00838 CPI (g/day) based on the ad libitum intake of diets of two different protein concentrations. Daily water intake by Standardbred horses fed 291 g protein/100 kg BW from forage was 29% higher than by horses fed 200 g protein/100 kg BW also from forage (Connysson 2009). Yet in a recent study, we found that the longitudinal water intake by light and light-cross pregnant mares (31/group) fed either a 7 or 12% protein diet for a 20-week period did not differ (p = 0.96). Mares, weighing 603 kg, and eating 2.05 kg DM diet/100 kg BW drank 52.6 ml water/kg BW despite a difference in protein intake of 65% (Cymbaluk, unpublished data).
Temperature effects and water intake
Horses adapt to cold and hot ambient temperatures and changes in water intake are part of this adaptive process (Cymbaluk & Christison 1990, Geor et al 2000). Cold weather (below –8°C) reduced water intake by 6 to 14% by yearling horses with no concomitant reduction in feed intake (Cymbaluk 1990b). Water intakes increased from 23.4 liters/day at thermoneutrality (20°C, 45–50% RH) to 39.5 liters/day after 20 days of short-term exposure (4 h daily) to a hot, humid environment (33–35°C, 80–85% RH) (Geor et al 1996).
Water intake and pregnancy and lactation
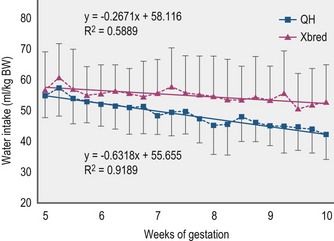
Pregnancy does not appear to impose an increase in water needs beyond maintenance. Pregnant, stabled mares eating 2.2 kg DM hay-grain diet/100 kg BW drank 45–69 ml water/kg BW (Freeman et al 1999, Houpt et al 2000). Weight-scaled water intakes by mares declined during pregnancy without a reduction in feed intake (Fig. 4.3). Absolute water intake remained steady despite an increased BW with advancing gestation.
Few data have quantified water intakes by lactating mares but water intake is felt to increase significantly above maintenance and pregnancy needs as a result of fluid loss (milk) and the increased feed intake associated with lactation (Doreau et al 1992). Pregnant mares fed grass hay ad libitum and less than 0.5 kg/100 kg BW concentrate ate 2.2 kg DM/100 kg BW daily (Freeman et al 1999) compared to lactating draft mares, which ate 3 kg DM /100 kg BW of diets high in forage or concentrate (Doreau et al 1992). Based on these differences in feed intake, water intake by lactating mares would increase 40% solely to compensate for a higher feed intake with an added 20 to 30 ml/kg BW of fluid or water to compensate for milk loss. Total water intake by lactating mares housed at thermoneutrality could increase to 75–100 ml/kg BW or 150 to 200% above water intakes observed in pregnancy. However, lactating mares are typically fed diets with more grain than when they are pregnant which might mitigate the increase in water intake expected with lactation because of the lower water: feed ratio of grain.
Water intake by suckling foals
Daily fluid intake by foals at 11, 25, 39 days of age was estimated at 15.1, 14.2 and 16.6 kg/day or about 255, 188 and 182 ml/kg BW (Oftedal et al 1983), values roughly fourfold greater than for mature horses. Orphan foals drank 148 to 159 ml fluid/kg BW (water plus milk replacer) at 1 week old then decreased fluid intake to 102 to 108 ml/kg BW by 7 weeks old (Cymbaluk et al 1993). Nursing and orphan foals begin drinking supplemental water in addition to dam’s milk or milk replacer at a relatively young age. Suckling 1-mo old foals drank 3.9 kg of water in addition to 17.4 kg milk and continued to increase water intake (5.5 kg/day) by 2-months old with no concurrent decrease in milk intake (Martin et al 1992).
Water or fluid intake in work or exercise
The impact of exercise on thermoregulation and fluid balance has recently been reviewed (Coenen 2005). Water needs of working or competitive horses are affected by a number of variables that influence sweat and respiratory fluid losses. The goal of fluid provision to exercised horses is to prevent dehydration and/or facilitate rehydration. Mild dehydration is described as a state of 5% total body fluid loss. In a 500-kg horse, whose total body water is about 350 liters, mild dehydration is a fluid loss of 17.5 liters whereas severely dehydrated horses have lost twice this amount. Not only is fluid lost, significant amounts of electrolytes are lost. Equine sweat is hypertonic to plasma (Lindinger 2008), while the dehydration created by sweating during vigorous exercise is hypotonic or isotonic. The plasma osmolality of horses with exercise-induced dehydration is insufficiently elevated to stimulate thirst (Waller et al 2008). Thus, providing only water, which has few electrolytes, to a horse in this physiological state will exacerbate plasma hypotonicity and further suppress the desire to drink. Strategies used for rehydration are discussed below.
Transportation
Transported horses exist in a temporary environment that can impose unique environmental, behavioral and dietary conditions. Horses transported for longer than 30 h without water became unfit for further transport whereas watered horses could tolerate an additional 2 h of transport (Friend 2000). Horses in transit consume water in response to thermal conditions, drinking more during hot temperatures and less during the cool periods of the day. During transport, both water and feed intake decrease, in amounts that are quite variable (Smith et al 1996). Light horses, accustomed to van transport, transported for 24 h during day time temperatures exceeding 30°C with humidity greater than 50%, lost 6% of body weight likely as fluid loss despite five stops to allow water consumption during the trip (Stull & Rodiek 2000). Weight losses of 3% were sustained in these horses for 24 h post-transport even though water was available ad libitum. Provision of water by bucket or trough should be considered for all transported horses and should be provided often (at least every 4 to 8 h) depending on weather conditions of the transport.
Drinking behavior of horses
True thirst or primary drinking results from a stimulus or desire caused by the need to replace fluid lost from intracellular and extracellular fluid reservoirs. Thirst is a response to changed plasma osmolality or blood volume, increased plasma sodium concentration or increased arterial blood pressure (Houpt & Yang 1995, Stricker & Sved 2000). Increases in plasma osmolality of 3% or less (about 8 mOsm) initiated drinking by horses (Sufit et al 1985, Jones et al 1989). Working horses in hot humid conditions that had high plasma osmolality (average 283 mOsm/kg) drank more frequently for longer drinking bouts and thus, had higher water intake than horses with lower plasma osmolality (Pritchard et al 2008). For idle horses, feed ingestion is the main reason for an increased plasma osmolality, which can increase by 3 to 5% within 1 h after feeding. Peri-prandial drinking (10 min before and 30 min after eating) is observed 75 to 89% of the time in horses fed ad libitum or four times daily (Sufit et al 1985
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


