Chapter 55 The Nutrition of “Browsers”
Historically, “browsers”—whether leaf-eating primates, browsing ruminants, or browsing rhinoceroses—have often been considered difficult to maintain under conventional zoo feeding regimens. Although epidemiologic studies are generally lacking, several problems seem to have a high prevalence in browsing species. Browsing ruminants seem to be particularly susceptible to rumen acidosis in captivity and also to “ill thrift” and poor body condition in general.11 In two large representative samples, the latter observation has led to explicitly recognized “syndromes”: the “wasting syndrome complex” in moose (Alces alces)9,69 and the “peracute mortality syndrome” or “serous fat atrophy syndrome” in giraffe (Giraffa camelopardalis).31,61 In tapirs (Tapirus spp.)48 and langurs,56 a soft fecal consistency is frequently observed, and browsing arboreal foregut fermenters, such as sloths or langurs, have a high prevalence of gastrointestinal (GI) upsets.25,26
BROWSERS ARE HERBIVORES FIRST
Vertebrates cannot digest plant cell walls auto-enzymatically. Therefore, herbivorous animals have to rely on the fermentation activity of symbiotic gut bacteria for the digestion of plant cell walls.68 These bacteria are contained in one or two major fermentation sites within the GI tract: in a foregut, a hindgut, or both.
The basic challenge in herbivore nutrition is to maintain a healthy, stable gut microflora in the herbivorous animal. The most common problem in herbivore nutritional management is a relative lack of plant cell wall material (fiber) and a relative oversupply of easily digestible and fermentable substrates (mostly soluble carbohydrates, e.g., sugar and starch; in extreme cases, perhaps protein). Such an oversupply will lead to direct disturbances of the microflora in foregut fermenters (comparable to rumen acidosis in domestic ruminants). In hindgut fermenters, these substrates are primarily absorbed from the small intestine before reaching the hindgut fermentation site, where they will only cause disturbances (comparable to cecal acidosis in domestic horses, the major cause of laminitis) if given in particular oversupply. This dualism may be illustrated with two primate groups: in captivity, with an oversupply of easily fermentable carbohydrates in conventional diets, langurs (foregut fermenters) have a history of digestive upsets and malnutrition,26 whereas lemurs (omnivores and some hindgut-fermenting herbivores) have a history of being obese.68 Similarly, among the ungulates, foregut-fermenting browsers such as giraffe or moose have a history of poor body condition in captivity,9,62 whereas hindgut fermenters, such as tapirs, rhinos, or elephants, are often overweight.1,60,75
In zoos, browsers have been traditionally recognized as animals with a difficult nutritional management. Partly, this reflects a basic problem in herbivore nutrition. As evolved omnivores, humans value easily digestible carbohydrates such as sugars and starches. We have a long history of cultivating grains and fruits for their respective starch and sugar content, and we intuitively want to include these items in the diets of animals we keep. Herbivores have also evolved to select for these items. In the natural environment, their sparse availability limits any potential danger of oversupply; in captivity, however, situations might exist in which the offer of these items is not limited. A restricted supply of such items, with a generous supply of high-fiber feeds, is therefore the fundamental approach to herbivore nutrition.
BROWSERS ARE DIFFERENT
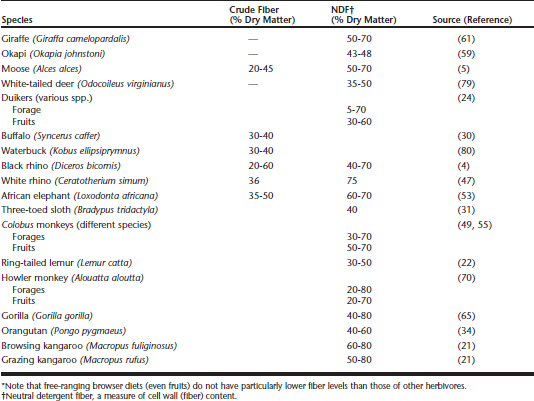
Nevertheless, compared with other herbivores, browsers still appear to be particularly sensitive. Possibly, one should not prioritize the investigation of other nutritional factors besides a high-fiber diet at first. In free-ranging browsers, a high fiber content is the major determining characteristic of their natural diet (Table 55-1); free-ranging browser diets are not distinctively “lower in fiber” than those of grazing species. In terms of fiber and carbohydrate content, a pelleted feed for “browsers” should therefore be suitable for “grazers” as well.
For the digestive physiology of many species, the provision of nonpelleted high-fiber diet items, that is, forage, is crucial. In ruminants, for example, the provision of structured fiber is a prerogative for proper rumen function, and in most herbivores, forage material is the only guarantee for a “normal” fecal consistency. In addition, forage addresses the most basic ethologic requirements of animals that have evolved physiologic and psychologic adaptations for the handling of (complex) food items, and oral stereotypies have been observed in many species such as cattle,64 okapi and giraffe,42 and horses81 eating forage-deprived or low-fiber diets. Particularly with respect to the suitability of forage, browsers are different: in contrast to the situation with grazing herbivores, the staple provision with a forage that is readily accepted by browsers without causing GI problems may be challenging in certain species.11
This general discrepancy between grazers and browsers is reflected in the traditional recipes for pelleted feeds provided by commercial manufacturers. Although there is no indication that browsers are adapted to a higher fiber intake than grazers, experience has led to the development of particular “browser” pellets high in fiber. In contrast, “regular herbivore” or “grazer” products contain less fiber because these animals readily accept the staple fiber source offered to them in addition to the pellets: grass hay. Using the product ranges of both U.S. Mazuri (PMI Nutrition International, St. Louis, Mo) and U.K. Mazuri (SDS, Essex),* it may be demonstrated that the recipes for ungulates increase in fiber content according to the difficulty of providing the target species with readily acceptable forage material, with moose being recognized as a large browsing species particularly reluctant to accept hay (Table 55-2).
Table 55-2 Declared Crude Fiber Content of Select Herbivore Feeds from Catalogs of Two Commercial Suppliers*
| Diet Name | Crude Fiber (% Dry Matter) | NDF† (% Dry Matter) |
|---|---|---|
| Herbivore 16-ADF1 | 16.7 | 32.2 |
| Herbivore 25-ADF1 | 25.6 | 43.4 |
| Browser breeder1 | 27.8 | 43.6 |
| Browser maintenance1 | 31.1 | 48.1 |
| Moose maintenance1 | 35.6 | 54.8 |
| Grazer2 | 11.2 | |
| Browser breeder2 | 18.6 | |
| Browser maintenance2 | 21.4 | |
| Moose2 | 24.0 |
* Note that fiber levels do not reflect differences in “fiber requirements” between target species, but that fiber content increases with the recognized target species’ reluctance to accept grass or alfalfa (lucerne) hay forage.
† Neutral detergent fiber, a measure of cell wall (fiber) content.
1 Mazuri (PMI, St Louis, Mo USA).
It should be noted that the fiber levels reported in Table 55-2 are still in the lower range of the fiber levels reported in diets of free-ranging animals (see Table 55-1). Interestingly, this discrepancy is even greater if the primate diets of commercial manufacturers (Table 55-3) are compared to the diets of free-ranging primates (see Table 55-1).
Table 55-3 Declared Crude Fiber Content of Select Primate Feeds
| Diet Name | Crude Fiber (% Dry Matter) | NDF* (% Dry Matter) |
|---|---|---|
| Leaf-Eater Primate diet1 | 15.6 | 27.4 |
| Primate High Fiber Sticks1 | 16.1 | 32.9 |
| Primate Browse Biscuit1 | 17.8 | 29.4 |
| Leaf Eater Primate2 | 13.8 | |
| High-fiber primate diet3 | 10.0 | |
| Leaf Eater Red Apple4 | 14.4 | 20.8 |
1 Mazuri (PMI, St Louis, Mo USA).
4 Marion Zoological (Plymouth, Minn USA).
DIFFERENCES BETWEEN GRAZERS AND BROWSERS
A fascinating multitude of anatomic and physiologic differences has been postulated (and sometimes demonstrated) between browsing and grazing herbivores and their respective diets.18 Internal and external differences in muzzle width, tooth form, salivary gland size and saliva composition, and GI tract morphology have been suggested as underlying features of importance in differential nutrition of browsing herbivores. Not all these differences are of direct relevance for zoo animal feeding. The following differences, however, most likely are important for the nutritional management of browsers:
Chemical Composition of Forages
Browse is regularly reported to contain more protein than grasses.18 On the one hand, this is most likely due to analytic difficulties: protein content is usually assessed by analyzing nitrogen and multiplying by the factor of 6.25. However, browse may contain significant amounts of nonproteinaceous nitrogen in secondary plant compounds, and it has been suggested, at least for tropical browse, that the true conversion factor for the calculation should be as low as 4.4.54 Lignin also contains nitrogen in a chemically unavailable form.77 Therefore, bound nitrogen may erroneously contribute to higher protein values reported for many browses unless available versus bound protein fractions are analyzed separately. On the other hand, high reported protein contents should not automatically lead to the assumption that browsers have higher protein requirements.
A classic case of an assumed high protein requirement in a browser is that of giraffe. It was suspected that low-protein diets play an important role in the serous fat atrophy syndrome (peracute mortality syndrome) observed in captive giraffes,32 and high protein levels of 18% dry matter (DM) were consistently recommended for this species. However, it was later reported that the problem also occurs in animals with “adequate” protein provision.45 A comparative evaluation of experimentally determined protein requirements in ruminants does not reveal relevant differences between the different feeding types.11 The fact that browsers do have higher fecal and urinary nitrogen losses when kept on browse does not reflect higher true endogenous losses, but rather is caused by the secondary plant compounds in the browse fed. If the same animals are kept on a diet without secondary compounds, the nitrogen balance is “back to normal.”66 As a logical consequence of such considerations, the recommended protein levels for giraffe, for example, have recently been reduced to 12% DM.34 Particularly high protein levels for browsers appear unnecessary.
Browsing ruminants have traditionally been termed “concentrate selectors.”40 This may have led to a widespread conception that browsers particularly prefer (and require) easily digestible carbohydrates such as starch and sugars, and that such animals may receive higher proportions of concentrate feeds. However, comparative evaluations of ruminant necropsies have revealed a higher prevalence of GI disorders, particularly acidosis, in browsers compared with grazers. Evidently, the so-called concentrate selectors often suffer from a condition triggered by too much “concentrate feeds.”11 Therefore, for didactic reasons alone, the term “concentrate selector” should be avoided. Actually, there is no indication that browse has a higher sugar or starch content than grass; however, browse contains higher proportions of soluble fibers, such as pectins.66 Pectin sources have been recognized in domestic ruminant nutrition as high-energy “concentrates” that may favorably replace starch-containing grain products because of a significantly less acidotic potential compared to grains.78 Therefore, pectin sources may be considered excellent energy-supplying diet items, both for browsers and grazers alike.41,46
The fiber component of browse is generally more lignified than that of grasses.77 Lignin is a basically indigestible material, and thus more highly lignified fiber is less digestible. Broad surveys of various forages have shown lignification indices (% lignin/% neutral detergent fiber [NDF]) approaching 20% to 30% of total fiber in browses, compared with perhaps half that proportion in grasses. To date, no observations indicate that a deliberate inclusion of lignin, rather than a general increase in overall fiber levels, is of particular health relevance for browsing animals. However, it is most certainly associated with decreased forage digestibility in a variety of herbivore species.
Browse plants, in particular the leaves of woody plants, often contain secondary plant compounds that may act as digestibility reducers or toxins that may serve as feeding deterrents. As adaptations, browsing animals may produce salivary proteins that reduce the effect of such substances (e.g., the tannin-binding proteins), and are likely to have evolved a variety of metabolic detoxification mechanisms.6 Because of the enormous variety of secondary plant compounds, general rules are difficult to distill from the literature. Some positive effects of some of these compounds have been reported (e.g., as antioxidants or anthelmintic substances, or by “protecting” dietary protein from ruminal degradation and thus enhancing intestinal digestion). In roe deer (Capreolus capreolus) fawns, an increased food conversion and a tendency for higher circulating antioxidant levels have been reported on a pelleted diet with added tannin.12 In black rhinoceros (Diceros bicornis) feces, the antioxidant capacity was higher on tannin-containing diets.17 However, the experimental evidence is still extremely limited. More studies are needed before any recommendations about the deliberate inclusion of such substances may be made.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


