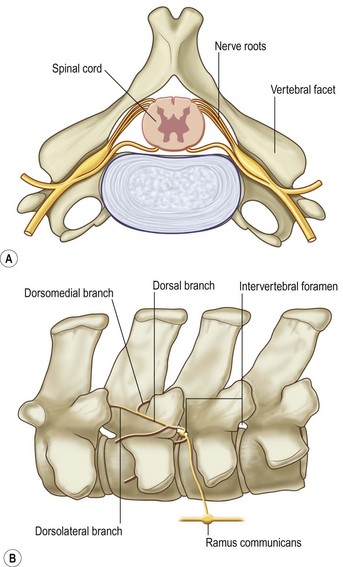
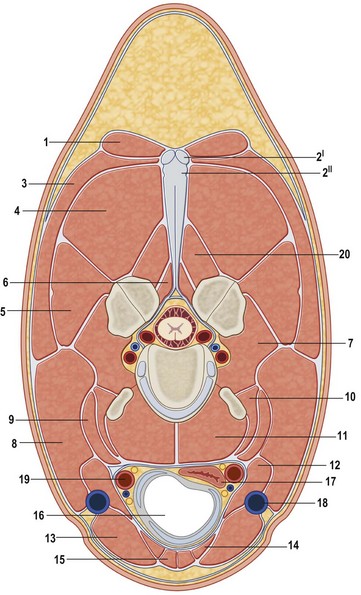
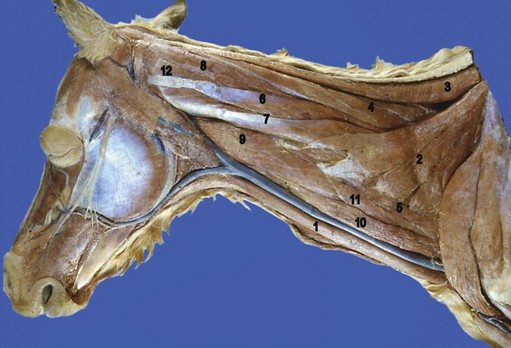
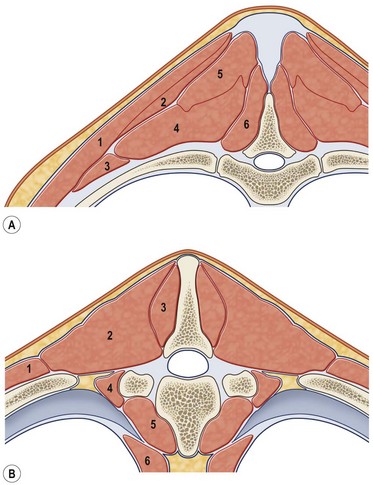
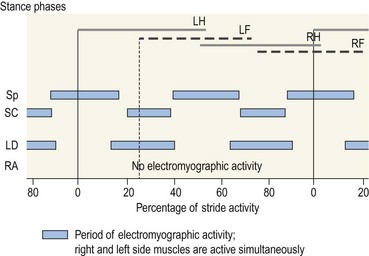
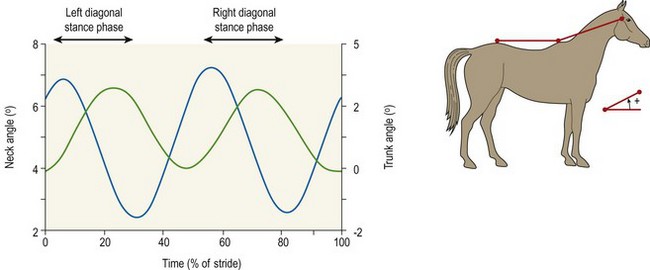
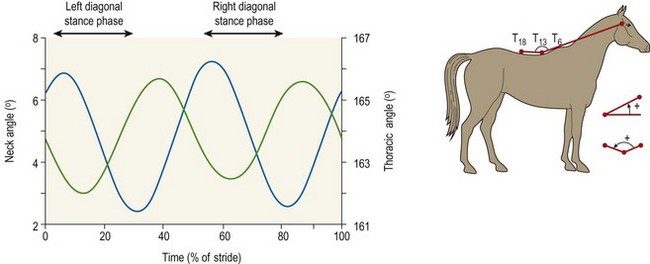
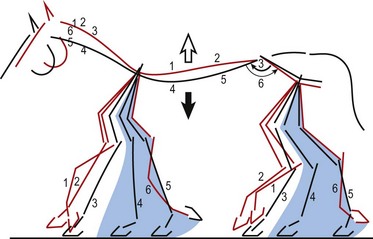
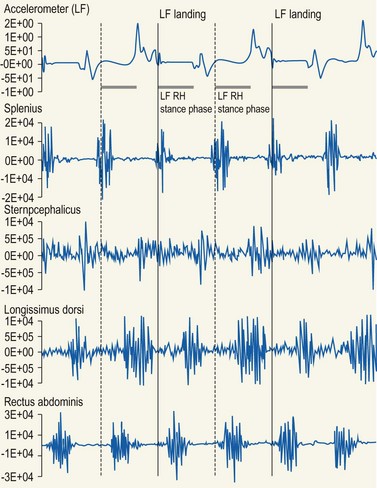
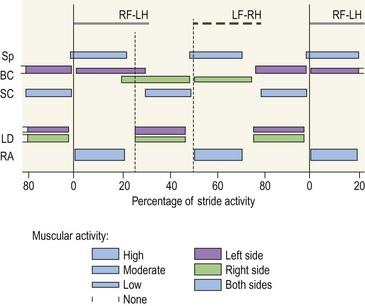
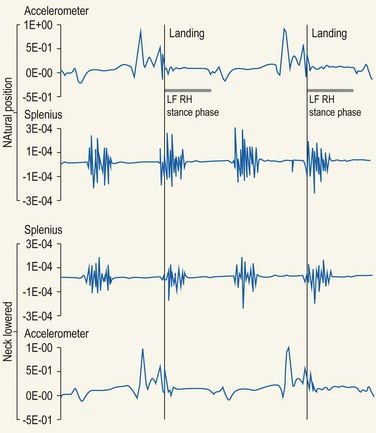
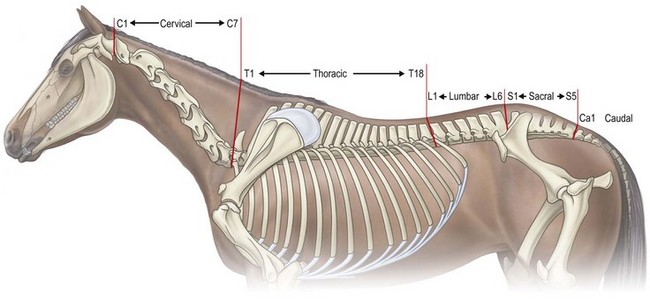
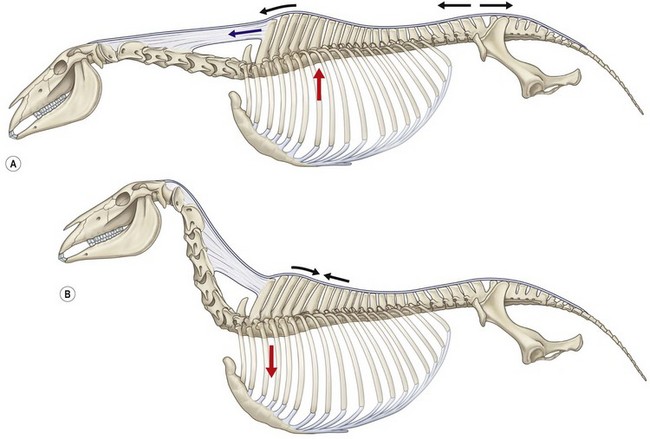
Chapter 10 Claudia Wolschrijn, Fabrice Audigié, Inge D. Wijnberg, Christopher Johnston, Jean-Marie Denoix and Willem Back The goal of this chapter is to present a synthesis of the current knowledge on in vivo neck and back function and dysfunction at walk, trot, gallop. Descriptions of equine vertebral column movements are often limited to the relative displacements of different axial regions of the horse (mainly neck versus back). These displacements induced and controlled by long muscle chains have been measured using kinematics and accelerometrics, of which in general surface EMG measures muscular activity and needle EMG muscular functionality. An anatomical overview of the vertebral column precedes the locomotor data review on its function and dysfunction (Clayton et al., 2005). The spinal column of the horse extends from the occipital condyles to the tail and consists of approximately 50 separate short bones, the vertebrae. The spinal column supports the body axis and thus the maintenance of posture. The vertebrae are connected by short and long ligaments, and two types of articulations, and therefore, the spinal column also plays an important role in locomotion. The typical vertebral formula for the horse is seven cervical, 18 thoracic, six lumbar, five sacral, and 15–21 caudal vertebrae (Fig. 10.1), each with its own specific characteristics. The basic form of vertebrae (this does not account for the first cervical vertebra) consists of a ventrally placed vertebral body and a dorsally placed vertebral arch, which encloses the vertebral foramen. The body is composed of an outer cortex of compact bone, which surrounds trabecular bone. The cranial aspect of the body is usually convex, the caudal aspect concave. Both are covered with hyaline cartilage and can as such be regarded as the non-ossified part of the epiphysis of the vertebral body. The basal part of the arch is notched on both the cranial and caudal margin; the two notches of successive vertebrae form one intervertebral foramen that transmits the spinal nerves and vessels (Fig. 10.2). The spinous process, which is located, on top of the arch, varies in its shape, length and inclination. On the left and right junctions of the body and the arch the transverse processes can be found. Mamillary processes, if present, are located between the cranial articular process and the transverse process; accessory processes are located between the caudal articular process and the transverse process. The head and neck unit plays an important role in balancing and in locomotion. Changing the position of the head and neck changes the center of gravity, which provides means for changing the speed of the gaits and even movement on the same position. The thoracolumbar part of the vertebral column is relatively stiff; it has to oppose the forces exerted by the relatively heavy abdominal organs and it has to transmit the propulsive forces brought about by the hind limbs. Fig 10.1 The vertebral formula for the horse is seven cervical, 18 thoracic, six lumbar, five sacral, and 15–21 caudal vertebrae. Denoix, J.M., Pailloux, J.P., 1996. Anatomy and basic biomechanical concepts. In: Physical Therapy and Massage for the Horse, with permission from Manson Publishing Ltd. The 18 (range 17–19) thoracic vertebrae have short bodies, low articular processes, closely fitting vertebral arches and long spinous processes. The latter increase rapidly in size with T2–T9 forming the basis of the withers. Normally, the tip of T5 or T6 forms the highest point of the withers. Thereafter a gradual decline in height of the spinous processes can be seen. The spinous processes of the first 14 thoracic vertebrae have a backward inclination, the 15th one has an upright spinous process, and the processes of the remaining thoracic vertebrae and of the loin or lumbar vertebrae have a forward inclination. Serial features are an appearance of an additional mamillary process as a projection from the transverse process and its gradual migration to join the cranial articular process and a change toward the end of the number of the thoracic vertebrae in the character of the articular facets from the cervical to the lumbar pattern (Fig. 10.1). There are sternal (10) and asternal (8) ribs, the neck of the ribs become gradually shorter caudally. Costal facets are present on both extremities of the vertebral body for articulation with the rib heads and on the stubby transverse processes for the rib tubercles. The articular facets of the heads and tubercle become confluent in the caudal thoracic vertebrae which increases the mobility of the last few pairs of ribs. The head of a rib has a cranial and a caudal facet, separated by a groove for the attachment of the intra-articular ligament of the head of the rib. The intervertebral discs between the vertebral bodies are composed of a pulpy nucleus and a fibrous ring (anulus fibrosus), although especially in the adult horse the distinction between these two parts is difficult (Yovich et al., 1985). The thickness of the discs decreases throughout the thoracic and lumbar region to reach their minimum thickness in the lumbar region. The pulpier nucleus is maintained under pressure and spreads the compressive forces over a wider surface. As a result the surrounding fibrous ring and the ventral and dorsal longitudinal ligaments are also brought under tension. Flexion of the vertebral arch brings about compression of the ventral side of the intervertebral disc and an upward (dorsal) movement of the disc. The structure and function of the nuchal ligament has been described by (Gellman & Bertram, 2002a, b). It supports the weight of the head, when the head is held high. The nuchal ligament is composed of two paired parts, the funiculus and the laminae. The funiculus originating from the external occipital protuberance receives the nuchal laminae at the level of the third cervical vertebra, and inserts on the spinous process of the fourth thoracic vertebra. The two parallel parts of the funiculus broaden in the region of the withers from where it continues as the supraspinous ligament to the caudal spinous processes of the lumbar vertebrae, and possibly the sacrum, until the first coccygeal vertebrae. The laminae (two sheets of tendinous tissue) originate from the spinous processes of the axis, the successive cervical vertebrae and insert on the first thoracic vertebrae. Between the nuchal ligament and the second or third thoracic vertebra a bursa (bursa subligamentosa supraspinalis or supraspinous bursa) is present. Additional bursae can be found between the nuchal ligament and the atlas or axis and are called bursa subligamentosa nuchalis cranialis and caudalis respectively. The fibrous supraspinous ligament is composed of two parallel parts like the funiculus of the nuchal ligament. In the lumbar region, the two parts can slope down from the top of the spinous processes. Its inner structure is built up of successive fascicles that span 3–4 vertebrae. In the thoracolumbar region the supraspinous ligament is partly fused with the thoracolumbar fascia to which the large extensor muscles of the back are attached. In the sacral region it unites with the gluteal fascia and the sacroiliac ligament. The supraspinous ligament plays an important role in biomechans of the back. It stretches with flexion of the vertebral column and relaxes upon extension; it thus opposes the spreading of the spinous processes (Fig. 10.3). Fig 10.3 The nuchal ligament supports the weight of the head, when the head is held high. It is composed of two paired parts, the funiculus and the laminae. Denoix, J.M., Pailloux, J.P., 1996. Anatomy and basic biomechanical concepts. In: Physical Therapy and Massage for the Horse, with permission from Manson Publishing Ltd. The forelimb is attached to the trunk through muscles, the so-called synsarcosis. The involved extrinsic muscles are the trapezius, rhomboid, pectoral (superficial and deep), and the ventral serrate muscle (Payne et al., 2004). The latter consists of a cervical part (longer fibers, less tendinous) (Fig. 10.4) and a thoracic part (short fibers, much tendinous tissue). The ventral serratus muscle is innervated by a brachial plexus nerve, the long thoracic nerve. The following four muscles are also extrinsic muscles and therefore might be considered to be part of the synsarcosis as well: the subclavius, the latissimus dorsi, the omotransverse and the brachiocephalic muscle. The synsarcosis muscles are all involved in stabilizing the position of the scapula relative to the trunk, but during movement they also assist in protraction or retraction of forelimb. The synsarcosis provides flexibility to absorb the impact forces upon landing on the ground. The trapezius muscle, consisting of a cervical and thoracic part, assists both in pro- and retraction (cervical, pulls the scapula to dorsocranial and thoracic part respectively). Both parts are motor innervated by the accessory nerve, the sensory innervation is transduced via the dorsal rami of the cervical and thoracic nerves. The rhomboid muscle lifts the scapula and turns the scapula to cranial. Dorsal branches of the cervical nerves and a special branch from the brachial plexus (dorsal nerve of the scapula) innervate the muscle (Fig. 10.5). The muscle covers the dorsoscapular ligament, which is unique in the horse. A second head of the rhomboid muscle (m. rhomboideus cervicis) (Fig. 10.6 (3)) runs cranially to the second cervical vertebra. The latissimus dorsi, which is a flexor of the shoulder joint, becomes a powerful retractor with aid of the ground reaction forces. The brachiocephalic muscle (cleidobrachialis, cleidocephalicus) is a protractor, but can also bend the neck and head or move it to one side upon action. It is closely related to the omotransversarius from which it is hard to separate. The cleidocephalicus and omotransversarius muscle are innervated by the accessory nerve, the cleidobrachial part by the axillary nerve. Sensory information is relayed by the ventral rami of the cervical nerves (Dyce et al., 2010) (Fig. 10.7). The following muscles coordinate the movements of the head, especially by the atlanto-occipital and atlantoaxial joints, like shaking, tilting, flexing and turning the head. The m. rectus capitis dorsalis major is located between the spine of the axis and the occiput–nuchal crest, lateral to the nuchal ligament; it contains a deep and superficial part. The muscle extends the atlanto–occipital joint and is innervated by the dorsal branch of the first cervical nerve. Beneath it lies the minor rectus capitus dorsalis muscle, which covers the dorsal atlanto–occipital membrane and shares the function and innervation with the major muscle. The lateral rectus capitis muscle is a small muscular band, which extends from the ventral arch of the atlas to the paracondylar process of the skull. It flexes the atlanto–occipital joint and tilts the head and is innervated by the ventral branch of the first cervical nerve. The ventral rectus capitis muscle between the ventral arch of the atlas and the base of the skull near the tympanic bulla also flexes the atlanto–occipital joint and has the same innervation as the lateral rectus muscle. The m. obliquus capitis cranialis extends obliquely craniolaterally over the atlanto–occipital joint, covered by the splenius and a part of the brachiocephalic muscle. It extends the atlanto–occipital joint and bends the head to the contracting side. It is innervated by the dorsal branch of the first cervical nerve. The m. obliquus capitis caudalis (Fig. 10.6 (12)) is positioned more caudally between the spinous process of the axis and the wing of the atlas; it receives its innervation through the dorsal branch of the second cervical vertebra. Unilateral action provokes rotation of the atlas and thus the head on the dens of the axis. Bilaterally it acts as a fixator of the head. A neck muscle, not belonging to the large extensor systems, is the splenius muscle, which is a large muscle, located at the dorsolateral part of the neck, under the cranial (cervical) parts of the rhomboid muscle. It originates from the cranial border of the thoracolumbar fascia, the spinal processes of the first three thoracic vertebrae and the median neck raphe. The cephalic parts inserts the nuchal crest and mastoid process with the longissimus capitis. The cervical part inserts on the caudal part of the alar wing and the transverse processes of third and fifth cervical vertebrae. The muscle is a neck extensor and upon unilateral action it moves the neck to the side. It is thought to play an important role in maintaining balance during galloping. The muscle is innervated by the dorsal branches of the cervical nerves. The long muscle of the head (m. longus capitis) (Fig. 10.6 (9)) is a strong muscle, which can be regarded as a cranial continuation of the long muscle of the neck. It has long muscles fibers, and is located on both sides next to the ventral part of the second to fourth cervical vertebra. It unites with the contralateral muscle and ends between the guttural pouches on the base of the skull. It flexes the atlanto–occipital joint, draws the head sideways, or the neck downward and is innervated by the ventral branches of the first four cervical nerves. The segmented long muscle of the neck (m. longus cervicis) extends from the atlas to T6. It runs craniomedially to insert on the bodies of the more cranial cervical vertebrae near the midline. The muscle flexes the neck and is innervated by ventral branches of both the intercostal as the cervical nerves dependent on the part of the muscle. Of the scalenus muscle, only the ventral (Fig. 10.6 (11)) and middle part are present. It takes its origin from the first rib, and is divided by the brachial plexus. It inserts on the transverse processes of the third to seventh cervical vertebrae (middle part) and the seventh cervical vertebra (ventral part). The ventral branches of the fifth to eighth cervical nerve and the first two thoracic nerves innervate both parts. It flexes the neck, draws it downwards and bends it sideways; it also supports inspiration. The sternocephalic muscle (Fig. 10.6 (1)) has a fusiform body and a long cranial insertion tendon. Dependent on uni- or bilateral action it flexes the head or provides lateroflexion. The muscle has motor innervation from a large branch of the accessory nerve, the sensory information is relayed by the ventral branch of the second cervical spinal nerve. The epaxial musculature of the neck and back can, apart from the above mentioned muscles, be divided into three main systems, the iliocostal, the longissimus and the transversospinal system, that are located more or less next to each other in the thoracolumbar region, where extensive fusion takes place. The most median muscle (transversospinal system) group is deep to the other two groups, the most lateral one (iliocostal system) is very small in the cervical region. They lie parallel on both sides of the spinous process, and if they are well-developed the supraspinous ligament can be regarded as the bottom of the groove between the muscles. All three systems are divided into a lumbar, thoracic, cervical and capital part on the basis of their topographical position and local presence. Unilateral contraction evokes lateroflexion of the spine. The lumbar part of the iliocostalis and longissimus system share the same origin at the iliac crest, and are in this area strongly fused together, and covered with a shiny aponeurosis. The thoracolumbar part of the longissimus is composed of series of overlapping fascicles, that are cranioventrally orientated and span several vertebral segments. The iliocostalis muscle runs from the intermuscular fascia near the iliac crest (hence the name), but also fascicles arise from the transverse processes of the lumbar vertebrae and the ribs to attach to more cranial segments with glistening small tendons on the caudal edges of ribs 1–15 and the transverse process of the last cervical vertebra. In the neck this muscle is weakly developed (Fig. 10.6 (5)). The longissimus system is topographically divided into a lumbar, thoracic, cervical, atlantic (Fig. 10.6 (7)) and capital part (Fig. 10.6 (6)). Its segmental origin is still reflected in the numerous individual attachments of its muscle bundles. The origin is at the ilium and sacrum, which have their insertion on the spinous and transverse processes of the lumbar and thoracic vertebrae. The thoracodorsal part is covered with a fascia. The cervical longissimus muscle is the cranial continuation of the thoracic part. It has a triangular shape and consists of four separate bundles that insert on the transverse processes of the caudal cervical vertebrae. The atlantal longissimus muscle (Fig. 10.6 (7)) extends from the last four cervical and first two thoracic vertebrae to the wing of the atlas. It partly fuses with the cranial part of the omotransversarius muscle. The capital longissimus muscle (Fig. 10.6 (6)) is a separate muscle, located medial to the cervical part and the splenius muscle, originating at the first thoracic vertebrae and extending to the mastoid process of the skull. At the level of the wings of the atlas, it is strongly fused with the splenius muscle. The corresponding dorsal branches of the spinal nerves innervate these muscle parts. The muscle extends the back, is active during the swing phase, and is also responsible for raising the body during rearing and kicking with both the hind limbs. When unilaterally contracted, it flexes the vertebral column laterally or is responsible for rotation of the head. The transversospinalis system is the most medial and deepest of the neck and back extensors. Separate bundles connect the spinous and transverse processes with subsequent vertebrae in the lumbar, thoracic and cervical regions. Within the transversospinalis system two separate groups of muscles can be distinguished. The first one is located directly over the skeleton occupying the space between the spinous processes, the vertebral arches and the transverse processes, and is named the spinal system. If the muscle bundles extend between the spinal and the transverse processes of adjacent vertebrae they belong to the transversospinal system. It is common to name both systems together as the transversospinalis system. The fiber orientation is caudoventrolateral to craniodorsomedial. The spinalis thoracis and cervicis muscle passes between the spinous processes of adjacent vertebrae; it forms a common muscle belly, which bridges several segments. The muscle is present between the last lumbar vertebra and the third cervical vertebra; its fibers are craniodorsally orientated. The muscle is innervated by the dorsal branches of the topographically corresponding nerves. The semispinalis capitis muscle is a strong muscle that is located in the triangle between the skull, the transverse processes of the cervical vertebrae and the nuchal ligament, and is covered by the longissimus and splenius muscles. It is composed of two parts, dorsomedially the biventer cervicis (Fig. 10.6 (4)) and ventrolaterally the complexus (Fig. 10.6 (8)) muscle. The biventer cervicis has its origin from the cranial thoracic vertebrae and its insertion on the skull. The muscle can be easily recognized because of the connective tissue inscriptions on the muscle belly, which show the multi-segmented origin of the muscle. The semispinalis capitis muscle raises the head when contracted bilaterally and flexes the head and neck to the lateral side when acting unilaterally and is innervated by the dorsal branches of the cervical nerves. The superficial cervical epaxial muscles have been described by Gellman et al. (2002c). Recently, the multifidous musculature has been the subject of study in the horse, because it atrophies in people with lower back pain (Stubbs et al., 2011). The muscle is composed of numerous short bundles, which extend from the articular or mamillary processes of the lumbar vertebrae, and from the transverse processes of the thoracic and cervical vertebrae to the spinous processes of the preceding vertebrae. The bundles can span up to five segments in the thoracic region. Cranially it unites with the cranial oblique muscle of the head, caudally with the musculature of the tail. In the neck five bundles are present at each vertebral level that are innervated by the dorsal branches of the corresponding cervical nerves. The attachment of the hind limb is much more stable, compared to the front limb, to optimize the transmission of the thrust generated by the hind limbs to the back. The hind limb is connected to the pelvis through the hip joint, the pelvis itself is connected with the rigid iliosacral joint to the spine. Extrinsic muscles are responsible for pro- and retraction of the limb: the gluteal muscles (superficial, medial, deep and piriform), the sartorius, the rectus femoris, the biceps femoris, the semitendinosus and the semimembranosus muscles. Several in vitro studies have been performed on the biomechanics of the neck and back (Jeffcott & Dalin, 1980; Townsend et al., 1983; Townsend & Leach 1984; Denoix, 1987; Clayton & Townsend, 1989a,b; Sleutjens et al., 2010), but our knowledge of these areas is still incomplete. In vitro studies were mainly performed to evaluate the nature and maximal amount of regional or intervertebral movements of flexion and extension, axial rotation and lateral bending (Gellman & Bertram, 2002a,b). With the different protocols used, wide displacements were studied (Denoix, 1999a), but mobility of the vertebral column rarely reaches its maximal limits during locomotion and sporting performance, although back pain due to pressure, tension and/or shearing on vertebral structures (Denoix, 1999b) can alter locomotion of sport and racehorses (Jeffcott et al., 1982; Denoix, 1998; Jeffcott, 1980). These studies on cadavers found that the lumbar part of the spinal column was very rigid, especially with respect to lateroflexion, with increasing mobility in forward direction. The same applied to axial rotation. It should be emphasized, however, that, although this work is of great value as it gives insight into the potential for movement of various parts of the equine spine, it does not represent reality, as there was obviously no influence of any of the active structures. Of special interest for the biomechanical function of the vertebral column is head and neck movement influence the movement of the back. If the head is lowered, the nuchal ligament will exert a pull on the withers and flex the spinal column. Vice versa, lifting of the head will extend the back (Fig. 10.3). Understanding of this concept is of great importance in the athletic training of sport horses. The question of how the function of the back can best be described dates back to Antiquity. Galenus (130–200 AD) developed the concept of the ‘vaulted roof’ in which the back and the upper part of the rib cage form a roof over the abdominal and thoracic cavities. A collapse of this roof would be prevented by the spinous processes. However, the fact that the spinous processes do not make contact in the normal situation makes this representation improbable. The next concept was developed by Bergmann in 1847 and further elaborated by Zschokke (1892). This concept implies the representation of the back by a bridge that is resting on four piers (the limbs). The upper ledger represents the supraspinal ligament and withstands tensional forces, the lower ledger, the vertebral bodies, is loaded under compression. The smaller girders between both ledgers represent the spinous processes and the ligaments in between these. Although this concept was generally adhered to until World War II, it contains a basic error in that such a bridge will not be loaded by tension dorsally and by compression ventrally, but just the other way round. It was the zoologist Slijper who, in 1946, after a meticulous study of the anatomical form of the vertebrae and especially of the inclination of the spinous processes in various species, came up with the model that is still holding today. His so-called bow-and-string concept not only takes into account the vertebral column and the limbs, but also the sternum and musculature of the ventral abdomen. In this concept the vertebral column is a bow that is held under intrinsic tension by the abdominal wall (Fig. 10.3). Considering the complete equine vertebral column, the movements of the neck and trunk and structural and functional description of the behavior of the axial muscles during different gaits and sport exercises has been published previously (Denoix, 1987; Denoix & Pailloux, 1996; Denoix, 1999a,b). At present, few in vivo studies have been published. A kinematic method to evaluate back flexibility on standing horses was presented (Licka & Peham, 1998). Neck and back motion and coordination are very different according to the gait and movement that are being performed and there is a need for better knowledge of the nature and amplitude of in vivo equine vertebral mobility. Neck and back flexibility and active mobility are essential to the accomplishment of sport exercises and are the subject of constant observation and concern for trainers and riders (Colborne et al., 2001; Cassiat et al., 2004). The behavior of the osteoarticular components of the equine thoracolumbar vertebral column mainly based on in vitro investigations has been performed (Denoix, 1999a). Three major movements take place in the equine intervertebral joints (Jeffcott, 1980; Townsend et al., 1983; Clayton & Townsend, 1989a; Denoix, 1999a): • Flexion (ventral bending inducing a dorsal convexity) and extension (dorsal bending inducing a ventral convexity) movements, occurring in the median plane, around a transverse axis • Lateral bending (lateroflexion) to the left and right sides, developed in the horizontal plane, around a dorsoventral axis • Left or right axial rotation (left or right deviation of a vertebral body with respect to the following one) around a longitudinal axis. Translational movements of minor amplitude take place in a vertical transverse plane: • Vertical shearing (dorsoventral translation) is associated with flexion and extension movements • Transverse shearing (left to right displacement) is associated with lateroflexion and rotation. Longitudinal compression and tension occur in some parts of the intervertebral joints for every major movement of the vertebral column. Kinematic and accelerometric studies of head and trunk movement at walk and trot on treadmills have been performed (Barrey et al., 1994; Buchner et al., 1996). At the walk, the vertical displacement, velocity and acceleration of the head, withers and tuber sacrale showed a sinusoidal pattern with two similar oscillations during each stride. The height of the withers and tuber sacrale was minimal at the beginning of the stance phase and maximal at about midstance of both thoracic and pelvic limbs (Buchner et al., 1996). The acceleration signal recorded over the sternum showed also two similar dorsoventral deviations corresponding to each half-stride (Barrey et al., 1994). A complementary analysis of the walk on a treadmill using three-dimensional accelerometers fixed to the front of a saddle (Galloux et al., 1994) showed that, at the walk, the amplitude of movement was higher in the vertical axis than in the transverse or longitudinal axes; rotation around the transverse axis (pitching motion) was higher than rotation around the longitudinal axis (rolling) and vertical axis (twisting or yaw). Furthermore, the twisting movement was greater in the walk than in the other gaits. Head movement adaptations associated with a supporting forelimb lameness induced by pressure on the hoof sole resembled those of the trot: the amplitude of the dorsoventral oscillation decreased during the stance phase of the lame limb and increased during the contralateral limb stance phase (Buchner et al., 1996). Similarly, the maximum acceleration amplitude over the sternum was reduced during the stance phase of the lame forelimb (Barrey et al., 1994). For induced hind limb lameness, elevation of the tuber sacrale was slightly reduced during the lame stance phase while head and withers movements were not significantly altered (Buchner et al., 1996). Surface electromyographic (EMG) activity of neck and trunk muscles was performed while horses walked on a treadmill or on hard surfaces, using skin-mounted surface electrodes (Fig. 10.8). It was found that the left and right splenius muscles act simultaneously before the landing of each forelimb (or during the second half of the opposite forelimb stance phase) to elevate the head and neck and facilitate forelimb protraction by the elongated brachiocephalicus muscle. The sternocephalicus muscles had reciprocal activity to splenius muscles and act before and during the first half of each forelimb stance phase, the longissimus dorsi muscles act during the intermediate part of each hind limb stance phase to facilitate propulsion (Peham et al., 2001a,b; Licka et al., 2009), and the rectus abdominis muscles do not show any significant EMG activity during the walk (Tokuriki et al., 1997; Robert et al., 1998), and this is correlated to the limited vertical acceleration of the abdominal visceral mass due to the lack of a suspension phase in this gait. A previous study performed with intramuscular fine-wire electrodes on four horses with and without a rider reported that the multifidus lumborum muscle was active in the intermediate part of the stance phase of the ipsilateral hind limb and the obliquus externus abdominis muscle had intermittent activity (Tokuriki et al., 1991). On the standing horse, in the neck region, the splenius muscle showed some activity, while the sternocephalicus and brachiocephalicus muscles were silent (Tokuriki & Aoki, 1991). Clinical observation shows that it is probably during walking that the mobility of the thoracolumbar vertebral column is most diversified, with association of rotation and lateroflexion combined with limited movements of flexion and extension. Kinematic and electromyographic data of neck and trunk motion and coordination have been established on sound horses with a three-dimensional kinematic analysis system allowing simultaneous recording of the left and right sides of the horse (Audigié et al., 1999; Robert et al., 1998; Pourcelot, 1999). Lame horses have also been investigated using kinematic analysis (Buchner et al., 1996; Audigié et al., 1999). The vertical displacement versus time of the head, withers and tuber sacrale at trot shows a sinusoidal pattern (Buchner et al., 1996) with two symmetrical oscillations during a stride. The general orientation of the neck and trunk and their alignment vary during the stride (Fig. 10.9). With respect to the tuber sacrale position, the withers elevate during the first and intermediate parts of each diagonal stance phase (upward rotation of the trunk) and descend during the last part of the stance phase as well as during the suspension phase (downward rotation of the trunk). Additionally, the croup presents wider dorsoventral movements (lowering and elevation) than the withers. In sound trotting horses, the head position is highest in the first half of each diagonal stance phase (Vorstenbosch et al., 1997). During the major part of the stance phase (Fig. 10.9), the neck rotates downward and becomes closer to the horizontal, with the head reaching its lowest point before the end of this phase. During the suspension and beginning of the following stance phase the neck becomes more oblique (Fig. 10.9), and the head is elevated again (upward rotation of the neck). In early stance both the neck and trunk undergo an upward rotation; during the middle part of the stance phase the angle between the neck and trunk flexes (Fig. 10.9) and during the last part of the stance phase, the neck and trunk rotate downward to become closer to the horizontal. During the suspension phase, extension between the trunk and neck occurs (Fig. 10.9). A three-dimensional in vivo kinematic study of flexion and extension movements of the thoracolumbar spine was performed at the trot in 13 sound horses using five skin markers placed on the median plane of the back over the 6th and 13th thoracic spinal processes as well as at the thoracolumbar, lumbosacral and sacrocaudal junctions (Pourcelot et al., 1998; Audigié et al., 1999). This study showed that maximal thoracic extension occurs near midstance (Fig. 10.10). This passive movement is produced by the visceral mass inertia. The maximal thoracolumbar extension occurs in the second half of each stance phase, followed by the maximal lumbosacral extension at the end of the stance phase (Audigié et al., 1999). The maximal thoracic flexion takes place during the swing phase and is concomitant with an elevation of the neck (Figs 10.10, 10.11). It is followed by the maximal thoracolumbar flexion. Finally, the maximal flexion of the lumbosacral junction occurs at the end of the swing phase at the time of maximal protraction of the hind limb (Audigié et al., 1999). The influence of the neck orientation on the thoracic spine dorsoventral mobility has been investigated in cadaver specimens (Denoix, 1987). The results showed that elevation of the neck facilitates thoracic flexion (Fig. 10.11). An accelerometric device fixed over the sternum area by an elastic girth (Barrey et al., 1994) showed that, for horses trotting on a treadmill, the dorsoventral acceleration curves had a deviation corresponding to each half-stride. The height of the dorsoventral acceleration signal was linearly correlated with speed. Longitudinal accelerations were less repetitive than dorsoventral ones; their magnitude increased with the increasing speed of the gait. Analysis of the trot on a treadmill using three-dimensional accelerometers fixed to the saddle (Galloux et al., 1994) showed that the linear movements along the three axes (longitudinal, transverse and vertical) were similar in amplitude. Rotation around the transverse axis (pitching motion) was smaller in comparison with the other two gaits (walk and canter). The surface electromyographic activity of neck and trunk muscles at trot has been investigated on hard track surfaces, with or without riders, as well as on a treadmill (Tokuriki & Aoki, 1991; Tokuriki et al., 1991, 1997; Robert et al., 1998, 1999, 2001, 2002; Licka et al., 2004). Comparison of neck muscle activity and neck orientation demonstrates that the splenius muscle acts before and during the first part of the stance phase of each forelimb (Figs 10.12, 10.13) to limit lowering of the neck (antigravitational activity). As mentioned by Tokuriki and Aoki (1991), muscle activity tends to be higher during the stance phase of the contralateral forelimb than for the ipsilateral one (Fig. 10.12). The sternocephalicus muscles have a reciprocal activity during the suspension phase to control neck elevation (Figs 10.12, 10.13). The brachiocephalicus muscle activity is high during the later part of the ipsilateral forelimb stance phase and during the suspension phase (Fig. 10.13) to achieve protraction of the ipsilateral forelimb; this muscle is inactive during most of the stance phase of the ipsilateral forelimb in order to avoid limiting forelimb propulsion. Lowering of the neck using the reins induces a reduction of splenius muscle activity during the first half of the stance phase of each diagonal (Fig. 10.14). These data can be explained by the lower amplitude of the neck displacement as well as by increased tension in passive anatomical structures that support the head and neck, such as the nuchal ligament. Comparison of trunk muscle activity and thoracolumbar movements shows that the rectus abdominis muscle acts during the stance phase (Figs 10.12, 10.13) to limit the passive thoracolumbar extension induced by the visceral mass acceleration. The longissimus dorsi muscles act at the end of each stance phase and during the suspension phase (Figs 10.12, 10.13) to induce lumbosacral extension and facilitate hind limb propulsion as well as to stabilize the thoracolumbar spine as it flexes. A previous study (Tokuriki et al., 1991) showed that the multifidus lumborum and longissimus lumborum were active before and after lift-off of each hind limb, with the activity being higher for the ipsilateral hind limb. According to this study the obliquus externus abdominis and rectus abdominis had roughly reciprocal activity to these epiaxial muscles. In another study (Tokuriki et al., 1997), the longissimus lumborum was thought to play a role in limiting lateral bending of the trunk during symmetrical gaits. Dorsoventral movements of flexion and extension have been quantified in sound horses (Audigié et al., 1999; Pourcelot et al., 1998). The maximal range of vertical displacements occurred near the 13th thoracic vertebra and, with respect to the tuber sacrale, reached an average value of 1.5 ± 0.2 cm. In clinical cases with intervertebral osteoarthrosis a significant reduction of this range of motion was observed (Audigié et al., 1999). Kinematic analysis was performed in nine high-level vaulting horses cantering at 4–5 m/s on a circle. As no markers could be placed on the back, markers indicative of neck and trunk movements were placed over the zygomatic arch, shoulder joint and tuber coxae (Fig. 10.15). The head and neck were positioned with side reins and the horses were filmed under competitive conditions at a left lead canter from outside of the circle. The neck and trunk angle curves showed that the cranial part of the neck and trunk becomes lower during the intermediate part of the support phase (downward rotation) and elevates (upward rotation) during the leading forelimb stance phase and the suspension phase (Fig. 10.15). The orientation changes of the neck angle occur before those of the trunk. An analysis of the canter using three-dimensional accelerometers fixed to the saddle (Galloux et al., 1994) showed higher amplitudes of motion than for the other gaits (walk and trot), especially along the longitudinal and vertical axes. Rotation around the transverse axis (pitching motion) and around the longitudinal axis (rolling) were greater than in the other gaits, while twisting around the vertical axis was lower. EMG activity of trunk and neck muscles was recorded with surface electrodes at the canter (Figs 10.16, 10.17
The neck and back
Function
Anatomy
Bones
Intervertebral discs
Ligaments
Muscles and nerves
Biomechanics
Coordination and locomotion
Walk
Trot
Neck muscle activity and neck orientation
Effect of neck orientation on neck muscle activity
Trunk muscle activity and thoracolumbar movements
Canter
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The neck and back