CHAPTER 20 The Liver
TECHNIQUES (Box 20-1)
Various techniques for percutaneous sampling of the hepatobiliary system have been described.1–4 Complications following these procedures are rare and include hemorrhage and potential seeding of needle tracts by neoplastic cells. Because compromised hemostasis is a life-threatening concern in animals with liver disease, the risk for excessive bleeding should be assessed particularly in cats with prolonged anorexia and in severely ill patients.4,5 A thorough history should include questioning the owner about prior bleeding episodes and treatment with over-the-counter anticoagulant drugs, such as aspirin. This is followed by a complete physical examination and laboratory testing to evaluate platelet number (a platelet count or estimate from a blood smear), a buccal mucosal bleeding time (BMBT), and coagulation assays (prothrombin time, activated partial thromboplastin time, and possibly protein-induced vitamin k absence). Abnormal test results do not reliably predict a bleeding tendency, and hemorrhage can occur in animals with normal profiles.6 It is pragmatic to avoid aspirating the highly vascular liver if there are severe coagulation abnormalities and/or clinical signs of hemorrhage elsewhere in the body. If a liver aspirate is deemed an acceptable risk in a patient with mildly prolonged coagulation tests, it is prudent to perform it early in the day so that the hematocrit can be followed and interventional abdominal surgery performed if excess bleeding is detected.
Box 20-1 Sample Preparation and Submission Guidelines
Aspiration and needle biopsy are usually performed with the patient in dorsal or right lateral recumbency, but can be performed with the patient standing. Chemical restraint is unnecessary for many patients. With the patient in dorsal recumbency, a blind biopsy is performed by inserting (at a 30- to 45-degree angle to the skin in dogs, and almost vertically in cats) midway between the left costal arch and the end of the xiphoid process.1 If the animal is standing or lying on the right side, the collection site is found by percussing the intercostal spaces to locate the liver beneath the rib cage. The next intercostal space caudal to the space where the sound of percussion changes from resonant to dull is usually a satisfactory site for aspiration. The collection site can be determined by palpation if an enlarged liver extends beyond the rib cage. Increasingly, sensitive imaging procedures such as ultrasound are used to facilitate accurate sampling of the liver, including sites with normal and abnormal echogenecity.3,4
Once the needle is withdrawn, the syringe should be detached and filled with 1 to 2 ml of air. After reattaching the syringe to the needle, a small drop of sample (6 to 8 mm diameter) is expelled onto each of several glass slides. Place the drops near the center of the slides; sample that is smeared close to the end may not be stained by automated stainers and is difficult to evaluate microscopically. The sample is best spread gently into a smear by the squash method (see Chapter 1).
The liver can also be sampled by a nonaspiration technique, using a 26-gauge needle only, as described in Chapter 1. This method may reduce the likelihood of contaminating the specimen with excessive amounts of blood.
Smears should be dried quickly and protected from exposure to formalin fumes or liquid for optimal staining. Typically, if several smears are available, one or two are stained with a routine Romanowsky-type (blood) stain and then examined. Retain additional unstained smears so that special stains can be performed if indicated. A well-made squash smear of a highly cellular aspirate is shown (Figure 20-1).
NORMAL FINDINGS
Aspirates and impressions from normal livers consist largely of hepatocytes and variable amounts of peripheral blood. Hepatocytes exfoliate readily and are distributed in the smear as single cells and in cords and clusters. Normal hepatocytes (Figures 20-2 and 20-3) are large, round or slightly oval to polyhedral cells that have round nuclei and abundant cytoplasm. Slight variation in cell size is normal. Their cytoplasm contains many ribosomes, which stain blue, and other organelles that are unstained or slightly pink. Therefore, the overall color of the cytoplasm in most normal hepatocytes is light blue or lavender, with light pink cytoplasmic granules. The intensity of the color is variable and depends on the degree to which the cell has been spread. Hepatocytes in large clusters, which resist flattening, have a deeper blue cytoplasm than single cells or cells in flattened sheets. In older animals, lipofuscin pigment granules may be seen in the cytoplasm of the hepatocytes (see Pigments).
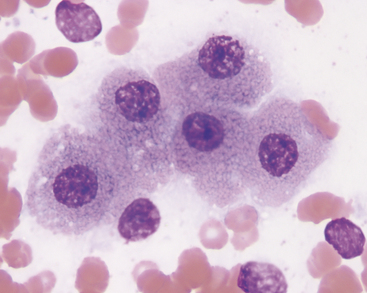
Nuclei typically are centrally located, are quite uniform in size, and have coarsely reticular chromatin. A single nucleolus often is visible; in some intact cells this can be obscured by heterochromatin. The nucleoli are more easily seen in the nuclei of ruptured or understained cells. A low number of binucleate hepatocytes is normal. Rectangular crystalline intranuclear inclusions are seen occasionally; these are of no known pathologic significance (see Figure 20-3).
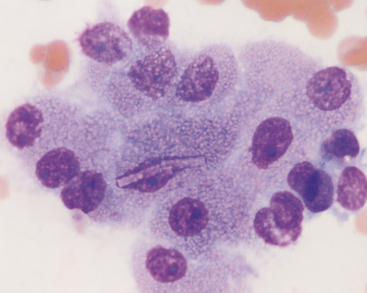
Biliary tract epithelial cells typically are few in aspirates of normal liver. These appear as cuboidal to columnar cells in small clusters or flat sheets (Figure 20-4). Rarely, distinct tubular arrangements may be seen (Figure 20-5). The cells have round, central nuclei and scant amounts of light blue cytoplasm, which distinguishes them from the larger, granular hepatocytes. Their chromatin is dense and smooth and nucleoli are not visible.
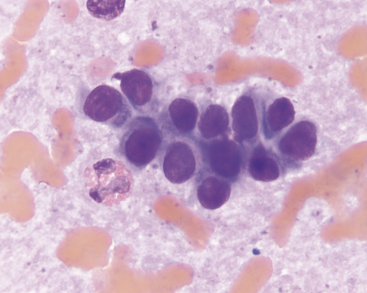
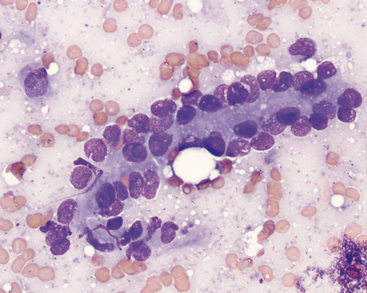
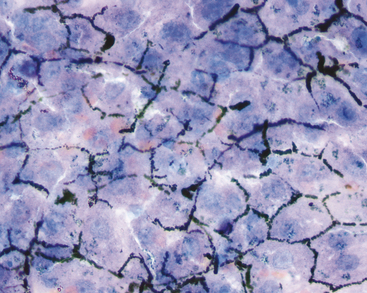
Mechanically exfoliated mesothelial cells may be seen in aspirates from some animals. These can be difficult to distinguish from biliary epithelial cells, but are generally larger, polygonal to spindloid, and found in flat sheets with a mosaic pattern (Figure 20-6). Occasional samples may include some amount of extrahepatic tissue such as adipocytes and/or lipid aspirated from mesenteric fat.
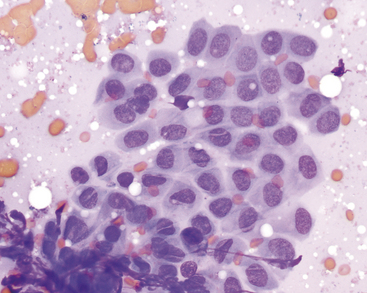
Resident hepatic macrophages, Kupffer cells, are few or absent in smears of normal liver. Rare lipid-laden Ito cells may be identified in some aspirates. In cats, low numbers of lymphocytes may also be seen and are of equivocal significance because a few lymphocytes can be found in the portal triads in healthy animals.7 Low numbers of resident mast cells are seen in many samples. These are small, often polygonal cells with a round blurry nucleus and lightly granulated cytoplasm (Figure 20-7). At lower magnifications, they will appear as small purple splotches nestled within the cords of hepatocytes.
PIGMENTS
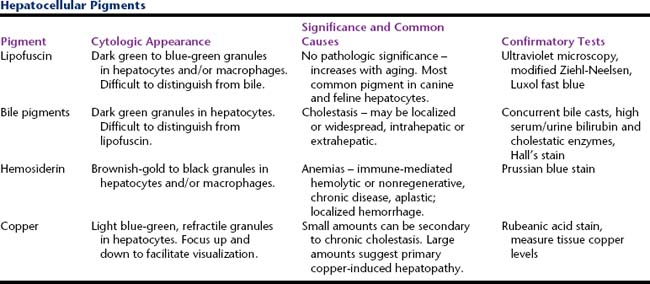
Several different pigments can be seen within hepatocytes in cytologic preparations (Table 20-1). Their presence can be a normal finding or be associated with extrahepatic or hepatic disease. Differentiating among these based solely on routinely stained smears can be difficult. Special stains and microscopy methods can be employed for specific identification. Typically this is necessary only when excess copper is suspected because this may be of direct diagnostic significance.
Lipofuscin
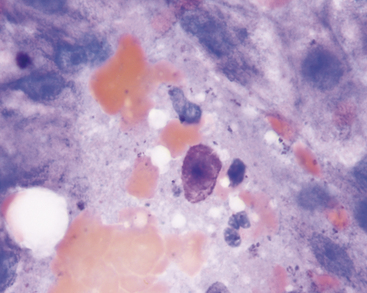
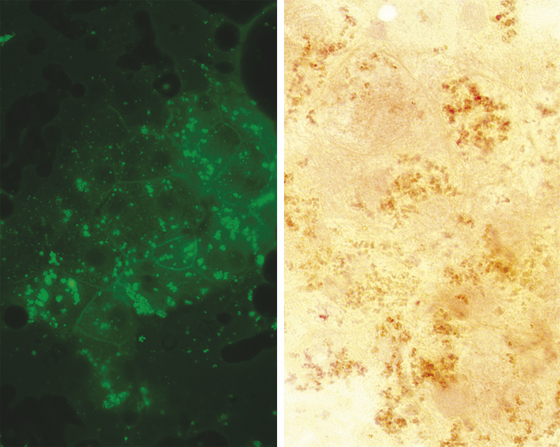
Lipofuscin is a “wear and tear” pigment, consisting of indigestible residue accumulated within autophagolysosomes. Large amounts may be seen in hepatocytes from older cats and dogs. This is a normal feature of aging and does not represent a pathologic process. Granules of lipofuscin (Figure 20-8) are similar in size to bile pigment and hemosiderin and range from brownish to blue-green in color. Based on one study, lipofuscin is the pigment most commonly observed within canine hepatocytes.8 The same is true in cats. Presumptive identification of hepatocyte pigment as lipofuscin, and not bile, is justified if other evidence of cholestasis (i.e., bile casts, hyperbilirubinemia, bilirubinuria) is absent. More direct identification of lipofuscin is seldom needed but can be achieved by examining unstained smears for autofluorescence under ultraviolet light at various wavelengths (Figure 20-9).8 Among the pigments discussed here, autofluorescence is specific for lipofuscin. Lipofuscin granules also specifically stain red with a modified Ziehl-Neelsen stain (Figure 20-9) or Luxol fast blue.
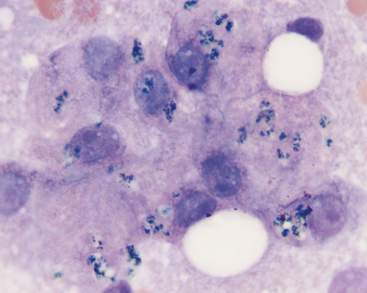
Figure 20-8 The dark pigment in these canine hepatocytes is lipofuscin. The cells were aspirated from a hyperplastic nodule in the liver of a 12-year-old dog. Images in Figure 20-9 are from the same aspirate. (Wright’s stain, 1000×.)
Bile Pigment
Bile accumulation within hepatocytes is recognized as small, dark green to black granules (Figure 20-10) and can be found in various hepatobiliary disorders. Abundant intracytoplasmic bile pigment is suggestive of cholestasis, although the process may not be severe enough to cause hyperphosphatasemia, hyperbilirubinemia, or icterus. Because of the similarity to lipofuscin with routine stains, however, cholestasis is confirmed only when solid, dark green or black bile casts, also called bile plugs or thrombi, are observed between contiguous hepatocytes (Figure 20-11). These represent accumulated bile in biliary canaliculi. Cast fragments may litter the background of the smears. In some cases, the cause of cholestasis may be evidenced by an accompanying inflammatory or neoplastic infiltrate.
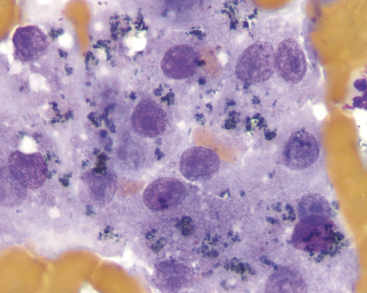
Figure 20-10 These hepatocytes contain a dark green to black granular pigment. Based solely on morphologic features, this could be either bile pigment (suggestive of cholestasis) or lipofuscin (no significance). In this instance, the concurrent findings of canalicular casts (see Figure 20-11) and serum biochemical evidence of cholestasis support the likelihood that it is bile. (Wright’s stain, 1000×.)
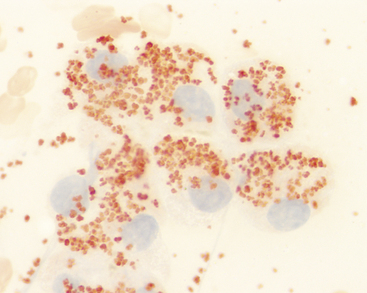
Hemosiderin
Hemosiderin is recognized as golden to golden-brown granules in hepatocytes (Figure 20-12). Hemosiderin can be distinguished from other pigments by applying a Prussian blue reaction to a methanol-fixed smear, which stains hemosiderin granules blue. In many cases, increased iron content in hepatocytes is associated with increased iron turnover caused by hemolytic anemia or ineffective erythropoiesis (e.g., immune-mediated nonregenerative anemia). Iron also can accumulate due to decreased iron utilization associated with severely decreased erythropoiesis (e.g., pure red cell aplasia, aplastic anemia). Repeated blood transfusions and administration of iron-containing compounds (especially by injection) also can increase hepatic iron content. In macrophages, hemosiderin can vary from gold to brown-black in color, depending on the amount within the cell. Macrophages containing phagocytized red cells or hemosiderin or both are concomitant findings in many patients with hemolytic disease.
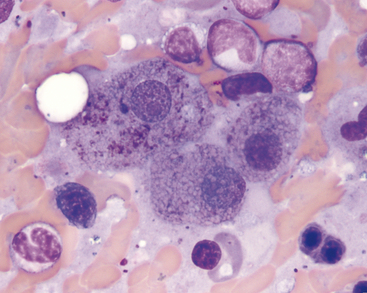
Copper
When present in excessive amounts, copper can become visible in hepatocytes as refractile, pale blue-green cytoplasmic granules (Figure 20-13). Smears can be stained with rubeanic acid to confirm the presence of copper (Figure 20-14). This procedure can be used even if the slide was first stained with Romanowsky-type stains. Small amounts of copper can accumulate secondary to prolonged cholestatic liver disease. If large amounts of copper are observed, a primary copper-accumulation hepatopathy should be suspected. This has been reported as a familial condition in Bedlington terriers, West Highland white terriers,9 Skye terriers,10 and Dalmations.11 In some breeds, such as the Doberman Pinscher, it remains unclear whether the increased copper represents a primary or secondary storage disorder.12 Copper-associated hepatopathy in a young Siamese cat has also been reported.13
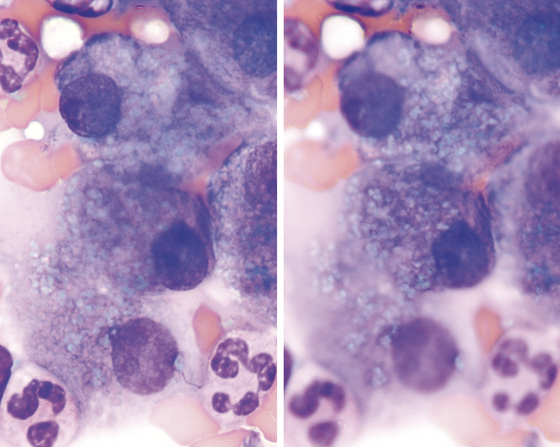
NONNEOPLASTIC CONDITIONS AND DISEASES
Hepatocellular Cytoplasmic Vacuolation
Morphologically, two types of cytoplasmic vacuolation are commonly seen: lipid and nonlipid (Table 20-2). Lipid vacuolation results from accumulation of triglycerides and is most often seen in aspirates of feline liver. Nonlipid vacuolation results from accumulation of glycogen and/or water, and is most often seen in aspirates of canine liver.

Lipid Vacuolation
The cytoplasm of affected hepatocytes contains round, empty-appearing, sharply delineated vacuoles of varying sizes (Figure 20-15). This appearance is termed fatty change or steatosis. The smear background may be punctuated with similar vacuoles. In severe cases, there are so many lipid vacuoles within the cells that little cytoplasm is visible. In some cells, a single very large vacuole may displace the nucleus to the periphery of the cell such that it resembles an adipocyte. The cells often are remarkably swollen by the lipid accumulation, which can contribute to cholestasis by compression of the canalicular spaces. Bile casts may be seen as a reflection of this effect. Lipid presence can be confirmed by staining an unfixed smear with Sudan III, Oil red O, or new methylene blue.
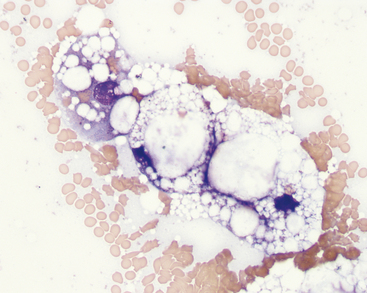
Fatty change of variable degree can be seen as a non-specific indicator of liver injury in all species. In cats, hepatomegaly due to fatty change is most commonly seen as the central feature in the cholestatic syndrome of feline hepatic lipidosis (FHL). Development of this disorder almost always is triggered by the presence of an underlying disease process causing a catabolic state.14 Affected cats are usually icteric and have livers that are enlarged by extensive, diffuse accumulation of lipid in hepatocytes. Most cats with hepatic lipidosis have markedly increased serum alkaline phosphatase activity and hyperbilirubinemia (mostly direct/conjugated) with no or only a slight increase in gamma glutamyltransferase activity. Hepatomegaly caused by lipid accumulation in hepatocytes is uncommon in dogs. Mild to moderate lipid vacuolation can develop secondary to various metabolic disorders in dogs and cats, especially diabetes mellitus. Severe hepatic lipidosis and hypoglycemia can occur subsequent to anorexia in puppies of toy dog breeds.15 Dogs with aflatoxicosis develop severe hepatic lipidosis; their liver aspirates resemble those of cats with FHL.
Nonlipid Vacuolation
The cytoplasm of affected hepatocytes has a lacey, wispy vacuolation or rarefaction often associated with mild to marked cell swelling (Figure 20-16). This appearance can be due to hepatocellular injury and has been attributed to increased cytoplasmic water. In dogs, a similar morphology can be due to glycogen accumulation, most notable and extensive in association with exogenous or endogenous glucocorticoid excess.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


