and Franca Placenza1
(1)
Department of Psychology, University of Toronto Scarborough, Toronto, ON, Canada
Abstract
The most insidious aspect of drug addiction in humans is a high and recurrent propensity to relapse. Over the past several decades, the reinstatement procedure has received widespread use as an animal model of drug relapse, to study the basic mechanisms underlying drug-seeking responses in laboratory animals. The objectives of this chapter are twofold. The first is to describe the primary paradigms and procedures that have been developed to study reinstatement of drug-related behaviors in the laboratory. The second is to define and characterize the three major triggers of reinstatement to drug seeking that constitute the foundation of this work. These triggers include priming injections of a previously self-administered drug, reexposure to drug-associated cues, and exposure to stress. The role of these triggers in reinstatement will be characterized within the context of an overview of key behavioral findings in the literature and their theoretical implications.
Key words
ReinstatementExtinctionSelf-administrationConditioned place preferenceConditioning1 Introduction
Drug addiction can be conceptualized as a chronic relapsing disorder, characterized by recurrent bouts of drug use with intervening periods of withdrawal and abstinence. Relapse may in fact represent the single-most predictive outcome of a diagnosis of addiction (1) and, as such, it poses one of the greatest challenges for treatment (2, 3). The problem of relapse is made more difficult by the fact that the time separating periods of drug use can vary on the order of hours and days to months and even years, and that different motivational influences may govern the likelihood of a full relapse episode depending on time since the last exposure to the drug (4–6). Furthermore, work carried out at a basic research level would suggest that the different triggers of relapse (e.g., a “taste” of the drug, reexposure to drug-associated cues, or exposure to stress) act via largely dissociable neuroanatomical systems to induce relapse (e.g., (7–13)) and may in fact be more or less likely to precipitate a relapse episode depending on the amount of time that follows termination of drug use (5, 14–18). From a treatment perspective, therefore, the problem of relapse is complex and requires careful consideration of multiple factors and influences.
Over the past several decades, a major research objective in the drug addiction field has been to understand the motivational processes and neurobiological underpinnings contributing to drug craving and relapse to drug use (see (5, 6, 12, 19–21) for recent reviews). In this regard, some of the major advances that have been made have come about as a result of studies carried out in the laboratory, using animal models of relapse. This work has focused in large part on identifying the triggers of relapse, and determining the conditions under which those triggers are most likely to precipitate a full relapse episode. In addition, a major emphasis has been on characterizing the neurobiological systems contributing to drug craving and relapse.
We will begin here by describing the major reinstatement procedures that have been developed over the past several decades to address mechanistic and phenomenological questions of relapse to drug seeking in the laboratory. Subsequently, an overview of some of the key behavioral findings and their theoretical implications will be provided, with an emphasis on studies aimed at characterizing the major triggers of reinstatement. These triggers include administration of a priming injection of a drug of experience, reexposure to drug-associated cues, and exposure to environmental or pharmacological stressors. Although, as mentioned, a major focus of drug reinstatement studies has been identifying the neurobiological systems involved, a review of these studies is beyond the scope of this chapter. The interested reader is, therefore, referred to several recent reviews on the topic (6, 7, 12, 22, 23).
2 The Reinstatement Model
Animal models of drug relapse share common features that qualify them as so-called reinstatement procedures. Thus, the term reinstatement is typically used to define relapse behavior in laboratory animals. Technically, reinstatement is defined as the resumption of a previously reinforced response when the unconditioned reinforcer or reward is presented non-contingently after a period of extinction training. During extinction training, the previously reinforced response gradually declines because reinforcement is withheld. Importantly, testing for reinstatement occurs under extinction conditions, in that responses are without scheduled consequence (24).
In addition to drug-seeking behaviors, reinstatement procedures have been used to study appetitive responses conditioned to food or sucrose (e.g., (25–28)), and aversive responses associated, primarily, with the conditioning of fear (e.g., (29–32)). Although the noncontingent presentation of the stimulus that originally maintained the behavior is what has traditionally defined reinstatement, these procedures have been adapted to study the ability of alternate stimuli, presented at test, to induce reinstatement. In fact, a major focus of the relevant drug literature in the past 10 years has been on environmental and pharmacological stressors as triggers for reinstatement (5, 33, 34). For example, acute exposure to intermittent footshock stress after a period of extinction training serves as a powerful stimulus for provoking the reinstatement of drug seeking in rats with a history of cocaine, heroin, nicotine, or alcohol self-administration (35–38).
From a historical perspective, the most conventional reinstatement procedures are based on the drug self-administration paradigm, in which animals are trained to perform an operant response for drug reinforcement (e.g., (24, 35, 39, 40)). More recently, alternate reinstatement procedures have been developed that are based on the conditioned place preference (CPP) paradigm, a widely used measure of the rewarding properties of drugs of abuse (e.g., (41–46)). In the next sections, these paradigms and their procedural variations for studying reinstatement will be described.
2.1 Reinstatement Procedures Based on the Drug Self-Administration Paradigm
A generalized schematic of the reinstatement procedure based on the drug self-administration paradigm is presented in Fig. 1. In reinstatement studies of this kind, animals are trained to perform an operant response, such as a lever press or nose poke, to obtain intravenous infusions of a psychostimulant (e.g., amphetamine, cocaine, nicotine) or opiate (e.g., heroin, morphine), or oral access to alcohol. Following training, extinction of the drug-reinforced behavior is accomplished by no longer reinforcing the behavior; that is, the operant response is either without consequence or results in an infusion of saline. Once the behavior is extinguished (i.e., responding is reduced to a low baseline level), animals are tested for the reinstatement of drug seeking induced by a specific triggering stimulus or event (e.g., priming injection of the drug, presentation of a drug-associated stimulus, or exposure to stress). Thus, the reinstatement of drug seeking is defined, operationally, as an increase above baseline in the number of occurrences of the previously reinforced behavior (e.g., number of lever presses on the previously reinforced lever) in response to the presentation of a triggering stimulus, after a period of extinction (24).
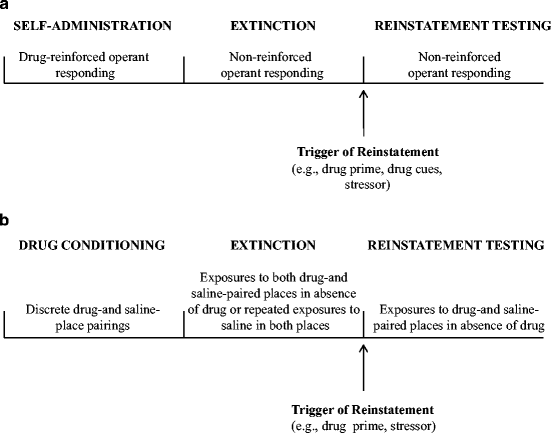
Fig. 1.
Generalized schematic of the phases of a reinstatement procedure based on: (a) the self-administration reinstatement procedure and (b) the conditioned place preference (CPP) procedure.
Since 1971, when the first report of drug-induced reinstatement of drug seeking appeared in the literature (40), three variations of the general procedure have been developed and used to study reinstatement of drug seeking in laboratory rats and monkeys. The differences that define these procedures relate primarily to whether drug self-administration, extinction, and testing sessions occur within the same day or on different days (47). Depending on the nature of the question being asked, some of these procedures are more suitable than others (see below), but reliable and consistent results have been obtained using all three procedures.
In one procedure, known as the “between-sessions” procedure (47), the various phases of the experiment occur sequentially, such that self-administration, extinction, and testing sessions are given on different days (e.g., (36, 40, 48)). Thus, animals are initially trained to self-administer a drug during consecutive daily sessions and, once stable self-administration is acquired (typically less than 20% variation from the mean number of infusions over several sessions), extinction conditions are introduced in subsequent daily sessions. After a predetermined extinction criterion is reached (e.g., 10 or fewer responses in a 3 h session or less than 20% responding on the last relative to first day of extinction), tests for reinstatement are given in the next daily sessions.
In an alternate procedure, the “within-sessions” procedure, self-administration training, extinction, and test sessions all occur within the same day (e.g., (39)). As in the case of the between-sessions procedure, animals are initially trained to self-administer a drug during daily sessions. In this case, however, once animals have acquired stable self-administration, subsequent daily sessions involve drug self-administration followed immediately by extinction training (to a predetermined extinction criterion) and then a test for reinstatement.
In the third procedure, the “between-within-sessions” procedure, once animals have acquired stable self-administration, sessions of extinction training and testing for reinstatement occur within the same day. Thus, drug self-administration occurs during daily sessions that are separate from extinction and testing; however, extinction and testing occur on the same day (e.g., (15, 49)). In studies using this procedure, animals are typically given multiple 1-hour daily extinction sessions separated by 5–30 min, in order to achieve stable, low levels of responding; these extinction sessions are then followed by a test for reinstatement. With the addition of a withdrawal period between the last day of self-administration training and the first day of extinction and testing for reinstatement, the “between-within” procedure has proved especially useful for studying the effects that different periods of drug withdrawal can have on extinction responding and reinstatement of drug seeking (15).
2.2 Procedures Based on the Conditioned Place Preference (CPP) Paradigm
In the past decade, several laboratories have adapted the CPP paradigm to study the reinstatement of extinguished preferences for previously drug-paired environments (41–46). CPP training is a Pavlovian conditioning procedure that involves discrete pairings of a drug, such as amphetamine, cocaine, or morphine, with a distinct environmental context, and pairings of saline with a different environmental context. At test, the animal is allowed to freely explore both contexts in a drug-free state. Evidence of a CPP is demonstrated by the animal spending more time in the drug- than saline-paired context, or relatively more time in the drug-paired context after than before the conditioning phase.
A generalized schematic of the CPP reinstatement procedure is presented in Fig. 1b. Two variations of the procedure have been developed that differ with respect to the way in which extinction is carried out. In one procedure, extinction is accomplished by giving repeated saline injections in both contexts, such that exposure to the previously drug-paired context is no longer rewarded (41–44). In the second procedure, animals are given repeated CPP tests on consecutive days. In this case, each test constitutes an extinction trial, since testing occurs in the absence of drug. In both procedures, testing for reinstatement is conducted by presenting the animals with a triggering stimulus, such as a noncontingent priming injection of the drug or exposure to an acute stressor, prior to providing the animal with free access to both the drug- and saline-paired context. Thus, reinstatement is defined operationally, as a proportionately greater amount of time spent in the previously drug-paired context, relative to saline-paired context, after a period of extinction.
3 The Triggers of Reinstatement
3.1 Drug Priming
One of the basic tenets of the organization Alcoholics Anonymous is that consumption of a single alcoholic beverage will greatly increase the probability of a full relapse. In fact, the idea that a small amount of drug can prime desire for more drug has been supported by several experimental findings obtained in psychostimulant and opiate-dependent subjects, and alcoholics (e.g., (50–56)). For example, in one study carried out with cocaine-experienced users, intravenous doses of the drug, relative to a placebo, were associated with marked increases in subjective ratings of cocaine craving (51). In another study, employing functional MRI, subjective ratings of cocaine-induced craving were closely correlated with focal signal increases in the nucleus accumbens, a key structure of the brain reward circuitry (50).
Studies of drug-priming-induced craving in experienced users parallel studies of drug-priming-induced reinstatement of drug seeking in rats and monkeys, in which noncontingent priming injections of a previously self-administered drug have been established as highly effective stimuli for inducing the reinstatement of drug seeking. In an original series of studies by Stretch and Gerber (40, 57, 58), noncontingent intravenous or intramuscular injections of amphetamine or cocaine produced a reliable reinstatement of responding in monkeys that had been trained to self-administer those drugs and had subsequently undergone extinction of the drug-taking behavior. In subsequent experiments using several variations of the reinstatement procedure, these effects were replicated in rats with histories of cocaine, heroin, and morphine self-administration (39, 59, 60). More recently, the drug priming effect was further extended to rats with histories of nicotine (61, 62), alcohol (37, 63), cannabinoid (64, 65), and MDMA (ecstasy) (66) self-administration. In addition, the drug priming effect was found to generalize to the CPP paradigm and, more specifically, the reinstatement of morphine- (43, 45, 67, 68) and cocaine- (44, 69) induced CPPs.
In addition to the drug of experience, drugs from the same pharmacological class as the self-administered drug can effectively induce reinstatement. For example, in monkeys trained to self-administer cocaine, priming injections of amphetamine induce the reinstatement of cocaine seeking (58, 70). Likewise, in rats, priming injections of amphetamine reinstate cocaine seeking (39), and priming injections of morphine reinstate heroin seeking (60). Drugs from different pharmacological classes than the self-administered drug, but affecting similar neurotransmitter systems, are also effective in inducing reinstatement of drug seeking. Such “crossover” priming effects have been demonstrated between opiates and psychostimulants (39, 60, 70, 71), nicotine and alcohol (72, 73), cannabinoids and psychostimulants (74), and cannabinoids and opiates (64, 65).
One explanation of the drug priming effect is that a noncontingent injection of the drug reinstates extinguished drug-seeking responses by activating the brain systems underlying its discriminative stimulus effects (40, 75). By this account, during testing for reinstatement, the priming injection elicits certain effects of the self-administered drug that signal to the animal the availability of response-contingent drug reinforcement. There are, however, several discrepancies between the outcomes of drug discrimination and drug priming studies that question the validity of this explanation. For example, some drugs shown to substitute for another drug in a drug discrimination procedure (e.g., D1-like receptor agonists for cocaine) do not induce reinstatement of responding for that drug (71). Conversely, some pharmacological manipulations that prime the reinstatement of drug seeking e.g., intra-VTA injections of morphine prime the reinstatement of heroin seeking (76) do not substitute for the drug in a drug discrimination procedure (35, 77). These findings suggest that the neurobiological systems underlying the discriminative stimulus and the priming effects of drugs on reinstatement are largely distinct. From this perspective, it is of interest that a neurobiological dissociation has also been reported between the discriminative stimulus and rewarding effects of drugs (77, 78), and that the priming effects of drugs have been attributed, at least in part, to the same brain systems that mediate their rewarding effects (79).
Indeed, findings from crossover priming studies with psychostimulants and opiates have led to the interpretation that activation of the mesocorticolimbic dopamine system, largely responsible for the rewarding effects of drugs of abuse, mediates reinstatement by both classes of drugs. For example, amphetamine injected into the nucleus accumbens reinstates heroin seeking (80) and injections of morphine into the ventral tegmental area reinstate cocaine seeking (76). Although these and similar findings are suggestive of a relationship between drug-priming-induced reinstatement and drug reward, other findings have failed to confirm a relationship. There are in fact several examples in the literature of pharmacological manipulations that either facilitate or interfere with drug self-administration, while having minimal or no effect on drug-priming-induced reinstatement of drug seeking. Likewise, there are examples of pharmacological manipulations that alter the effectiveness of drug-priming-induced reinstatement while having little or no effect on drug self-administration (see (23, 47)).
The mesocorticolimbic dopamine system is involved not only in the acute rewarding properties of drugs, but is also thought to underlie their incentive motivational and goal-directed effects (79, 81). As such, the reinstating effects of drug priming may also be associated with the ability of drugs to produce an incentive motivational state that leads to drug-seeking behavior (79, 81). According to this idea, priming injections of a drug enhance the incentive salience (i.e., “attractiveness”) of previously drug-related cues, and thus enhance behaviors that facilitate approach to those cues. Such cues may include a visual or auditory stimulus paired with an operant response (e.g., lever press) or, in the case of studies based on the CPP procedure, contextual cues. In support of this view, it has been shown that the removal of drug-associated cues during tests for reinstatement attenuates the effectiveness with which priming injections induce reinstatement of drug seeking (40).
3.2 Drug-Associated Cues
Clearly, events other than reexposure to drugs can provoke relapse to drug seeking. Indeed, after a period of abstinence, drug seeking is antecedent to drug taking. One such event that has been the focus of considerable study is exposure to cues that have become associated with the experience of drug taking. For example, reexposure to the people, places, or paraphenalia associated with drug use can elicit strong drug craving, and it has been argued that such cue-elicited craving is fundamental to the cycle of relapse in addiction (82). Given the crucial role that drug-associated cues are purported to play in drug craving and relapse to drug use, it is perhaps not surprising that reinstatement procedures have been used extensively to study the basic mechanisms mediating their effects.
In an initial laboratory demonstration of cue-induced reinstatement, Davis and Smith (1976) (59) trained rats to self-administer morphine under conditions in which a lever press response led to a drug infusion and the simultaneous sounding of a buzzer. Subsequently, animals underwent extinction training in the absence of the buzzer cue; that is, during extinction, lever presses led neither to drug infusions nor presentations of the buzzer. At the time of testing for reinstatement, animals were given a noncontingent presentation of the buzzer prior to access to the previously drug-reinforced lever. Under these conditions, reinstatement of responding was observed. In subsequent studies, the findings of Davis and Smith (59) were replicated and extended to animals trained to self-administer other drugs, including cocaine, heroin, ethanol, and nicotine, in combination with other types of discrete stimuli, such as a light cue or tone (39, 83–86).
Although the noncontingent presentation of a discrete drug-associated cue reliably reinstates drug seeking, it is generally not as effective in doing so as is, for example, a contingent presentation of a cue (39, 87) or a priming injection of the self-administered drug (e.g., (39)). This observation has led researchers to study factors that might influence the effectiveness with which a discrete cue induces reinstatement of drug seeking. The objective of this work, in part, has been to optimize the procedures for studying cue-induced reinstatement. For example, it has been shown that a compound drug-associated cue (e.g., light plus tone) is much more effective than a singular cue (light or tone) in inducing the reinstatement of drug seeking. In one study, response-contingent cocaine infusions led to varied presentations of a light, tone, or light plus tone compound stimulus (88). Following extinction training in the absence of the drug and cues, noncontingent presentation of the compound stimulus reinstated drug seeking, whereas presentations of either stimulus alone led to a much weaker effect on reinstatement, or was without effect. As mentioned previously, another factor that influences the effectiveness with which a drug cue induces reinstatement of drug seeking is its contingency of presentation at test. More specifically, it has been reported that presenting the discrete cue at test contingent on the operant response facilitates the reinstatement of drug seeking, relative to presenting the cue in a noncontingent fashion (39, 87).
In addition to discrete cues, discriminative cues that predict drug availability have been found to reinstate drug seeking. The first demonstration of this was obtained using a runway model of relapse, in which rats were initially trained to traverse a straight-arm maze for intravenous infusions of heroin in the goal box (89). On subsequent trials, the subjects learned to discriminate between two odors. One odor, the so-called S+, was accompanied by intravenous infusions of heroin in the goalbox and the other, the so-called S−, was accompanied by a saline infusion in the goalbox. Thus the S+ signaled the availability of heroin, whereas the S− signaled its absence. Discrimination training was considered to have occurred once the latency to reach the goalbox was less on S+ than S− trials, at which time extinction conditions were introduced. During extinction training, entries into the goalbox were without scheduled consequence. On test day, animals were given varied presentations of the S+ or S− before runway trials. Presentations of the S+, relative to the S−, reduced latencies to reach the goalbox; that is, the S+ reinstated the extinguished runway behavior.
More recently, the effect of discriminative cues on the reinstatement of drug seeking has been studied using the drug self-administration procedure (90–93). In these studies, rats are trained to perform an operant response (e.g., lever press) for intravenous infusions of a drug or saline, or for oral access to alcohol or water, during multiple daily sessions. Throughout sessions in which drug or alcohol is available, a discrete visual or auditory stimulus (i.e., S+) is presented; throughout sessions in which responding results in saline infusions or access to water, an alternate visual or auditory stimulus (i.e., S−) is presented. Following discrimination training and subsequent extinction sessions, in which responding is without scheduled consequence, noncontingent presentations of the S+, but not S−, are found to reinstate drug seeking.
Several studies have been carried out to characterize the phenomenology of reinstatement induced by discriminative cues. In one such study, rats that were trained to discriminate between the availability of intravenous infusions of cocaine versus the availability of saline exhibited strong reinstatement of cocaine seeking by presentations of the S+, throughout a month-long period of repeated testing, and following an additional 3-month drug-free period (92). These results suggest that cocaine-associated discriminative cues are highly resistant to extinction and are persistent in their motivational effects on drug-seeking responses. In another study in which rats were trained to discriminate between stimuli associated with the availability of alcohol versus water, reinstatement of alcohol seeking was induced by an olfactory, but not an auditory, discriminative stimulus. These results suggest that, at least in the case of alcohol-trained animals, the efficacy of discriminative stimuli in inducing reinstatement may depend on their modality of presentation (94).
Using an adapted so-called renewal procedure (32), Crombag and Shaham (2002) (95) demonstrated that the contextual cues associated with drug taking can also contribute to the reinstatement of extinguished drug-seeking responses. The renewal effect refers to the recovery of a conditioned response when testing occurs in a different context than that in which the response was extinguished. In the study by Crombag and Shaham (95), rats were trained to self-administer a heroin–cocaine speedball mixture in a context containing a set of distinctive visual, tactile, auditory, and olfactory stimuli. They subsequently underwent extinction training in an alternate context, containing a different set of distinctive visual, tactile, auditory, and olfactory stimuli. At the time of testing, animals that were returned to the context in which the original training had occurred exhibited renewed responding on the previously drug-reinforced lever.
Theoretical explanations for how drug-associated stimuli, be they discrete drug-associated cues, discriminative stimuli signaling the availability of drug, or contextual cues associated with the drug-taking environment, can come to acquire control over drug-seeking behaviors have been the focus of numerous seminal papers in the drug addiction field. Explanations have focused largely on Pavlovian learning processes, the idea of occasion setting, and the idea that drug-associated cues, through associative learning, can come to acquire incentive motivational properties that facilitate drug seeking. A comprehensive review of these varying accounts is beyond the scope of this chapter. The reader is therefore, referred to several relevant theoretical papers and recent reviews (33, 96–99).
3.3 Stress
There are many correlational studies implicating life stress as an important factor contributing to increased rates of drug and alcohol use, and as a trigger for relapse in individuals with substance abuse disorders (55, 100–104). A relationship between stress and relapse in drug addicts has also been studied under controlled laboratory conditions. For example, the induction of psychological stress, using a guided imagery procedure involving recall of personalized stress situations, was found to increase subjective reports of cocaine and alcohol craving in recently abstinent cocaine-dependent subjects seeking treatment (105, 106). Moreover, using these procedures, stress-induced cocaine craving reportedly predicted the incidence of cocaine relapse following inpatient treatment (see (105)).
Since 1995, reinstatement procedures have been used extensively to explore the relationship between stress and relapse to drug seeking in laboratory animals. To date, much of this work has used intermittent exposure to mild, electric footshocks as the stress manipulation. In an initial study carried out in animals trained to self-administer heroin, Shaham and Stewart (1995) (35) demonstrated that 10 min of exposure to brief intermittent electric footshocks served as a powerful stimulus for inducing the reinstatement of extinguished drug-seeking behavior. Furthermore, this effect of footshock on reinstatement persisted after an additional 4–6 week drug-free period and additional extinction training. In subsequent experiments, the effect of footshock on the reinstatement of heroin seeking was replicated and extended to animals with histories of cocaine, alcohol, and nicotine self-administration, as well as self-administration of a cocaine–heroin (“speedball”) mixture (see (6, 7, 23, 37, 38, 47)). The effect of footshock on the reinstatement of drug seeking was also found to generalize to the CPP paradigm, where it has been found to effectively induce reinstatement of both morphine and cocaine CPPs after drug-free periods of more than 1 month (42, 46, 107).
In addition to generalizing to the CPP procedure, the effects of footshock on reinstatement of drug seeking have been found to generalize to certain other types of stressors. One such highly effective stressor is 21 h of food deprivation, which induces the reinstatement of cocaine and heroin seeking at levels comparable to that induced by footshock stress (108–110). Likewise, various pharmacological stressors have been found to induce the reinstatement of drug seeking. For example, i.c.v. injections of the stress-related neuropeptide, corticotropin-releasing factor (CRF), reinstate heroin (111), cocaine (49), and alcohol (112) seeking, and injections of CRF into the bed nucleus of the stria terminalis (BNST), a brain region critically involved in the effects of stress on reinstatement, reinstate cocaine seeking (113). Likewise, systemic injections of the corticosterone synthesis inhibitor metyrapone, reinstate heroin seeking, presumably by acting acutely to reduce negative feedback in the hypothalamus and pituitary (111). Finally, central injections of the alpha-2 antagonist, yohimbine, reinstate responding in animals trained to self-administer alcohol (114), methamphetamine (115), or cocaine (116).
< div class='tao-gold-member'>
Only gold members can continue reading. Log In or Register to continue
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


