Chapter 33 Nutritional Factors Affecting Semen Quality in Felids
Proper nutrition is being increasingly recognized as a critical component of captive breeding programs for nondomestic cats.1 In many cases, especially when commercial feline diets are not available, the lack of reproduction may serve as a sensitive indicator of nutritional deficiencies and provide an early warning for the development of diet-related pathologic conditions. If nutritional problems are not addressed, conservation and breeding programs may fail to achieve their tremendous potential.
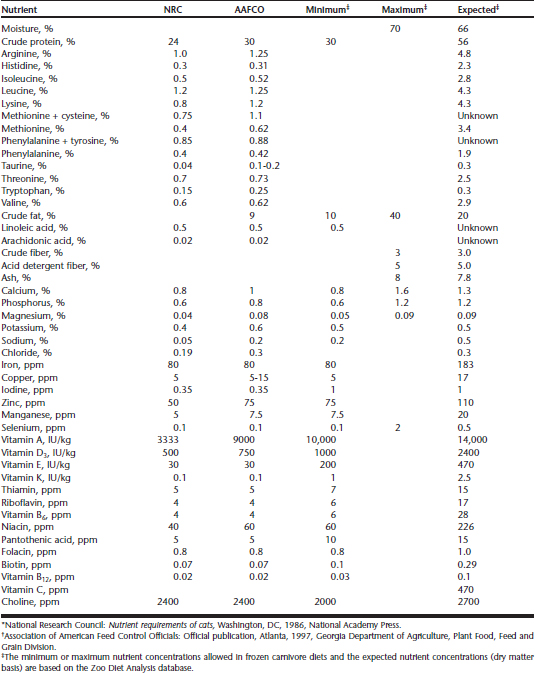
There is limited information on the nutrient requirements of most nondomestic felid species. Some digestibility studies have been conducted in small felids, such as the serval (Leptailurus serval), lynx (Lynx lynx), caracal (Caracal caracal), and sand cat (Felis margarita),6,10 and in large felids, including the tiger (Panthera tigris), lion (Panthera leo), puma (Puma concolor), and leopard (Panthera pardus).9,29,31 Although diets may be digested differently,15 the type of diet offered to captive nondomestic felids is based on nutritional requirements for domestic cats Table 33-1.3,19 Until more descriptive research is conducted regarding the nutrient requirements of nondomestic felids, extrapolation from domestic cats is necessary. In North American zoos, this strategy of using the domestic cat as a model for nutrition in the 36 species of nondomestic cats has been effective.
Table 33-1 Minimum NRC* Nutrient Concentrations Required in Purified Diets for the Growing Domestic Cat Compared with AAFCO† Nutrient Profiles for Growth and Reproduction of Cats Fed Practical Diets
PROTEIN AND AMINO ACIDS
The protein requirement of the cat is higher than that of most mammalian species studied.7 Generally, a protein requirement is actually a requirement for individual amino acids. The cat’s higher protein requirement may result from a need for more total protein, not only an increased requirement for essential amino acids.23 In general, proteins from animal matter contain a more balanced amino acid profile and better digestibility than plant proteins. However, the perfectly balanced protein complete in all essential amino acids has not been found for cats. Even the amino acid deficiencies of beef are evident when compared to the nutrient requirements of domestic cats.7 Amino acid availability also may be influenced by storage and food processing. Long-term storage may cause degradation of some nutrients. Certain amino acids may be either destroyed or rendered unavailable by the heating that often occurs during canning or extrusion processes.
Two amino acids have a special significance for cats, arginine and taurine. The cat is unusual in its reliance on the amino acid arginine. The cat with an arginine deficiency is unable to metabolize nitrogen compounds (through the urea cycle), which produces rapid elevation of blood ammonia levels resulting in ammonia toxicity and death.18 Other species may require arginine for growth, but in general they do not need it for adult maintenance.
Taurine also is an essential amino acid for cats. The particular importance of taurine in cat nutrition has been studied for more than 20 years. Cats depend on taurine for the formation of bile salts and cannot synthesize sufficient taurine to meet their needs. Taurine deficiency is linked with dilated cardiomyopathy and retinal degeneration.5,20,21,23 In leopard cats (Prionailurus bengalensis), taurine deficiency was observed in males and females fed a commercial canned cat food for 10 to 24 months, resulting in retinal degeneration ranging from focal tapetal lesions to diffuse pigment atrophy and blindness.12 Composition of the canned diet was modified to prevent dietary deficiencies. Interestingly, taurine deficiency and retinopathy had no influence on male reproductive function in these leopard cats. High concentrations of normal motile spermatozoa were detected each month during the diet-induced taurine deficiency.12
In female domestic cats, taurine depletion severely compromises reproductive performance, including an increase in fetal resorption, abortion, and stillbirth.26 Live-born kittens demonstrate numerous neurologic abnormalities, low birth weight, and poor postnatal survival rate caused by inadequate maternal lactation.26 The domestic cat’s minimum taurine requirement was studied,5,23 and a taurine content of 500 mg/kg dry matter of the diet was adequate for pregnancy in cats.19 However, when fed commercial canned diets, 2000 mg taurine/kg dry matter was needed.8,21 The cause may have been reduced gastrointestinal absorption of taurine or excessive excretion from the digestive tract. New studies suggest that heat treating of cat food may bind the free taurine, making it unavailable to the animal.20
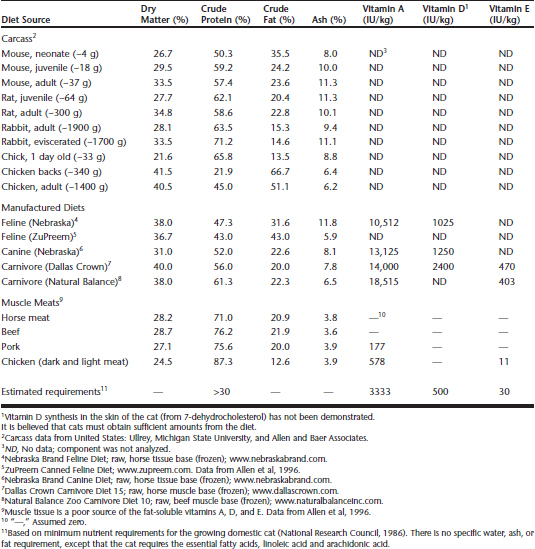
Table 33-2 lists the protein concentrations in “whole-prey” carcasses, with several common commercial diets and various types of muscle meats. An exclusive whole-prey diet consisting of an intact carcass containing bones and viscera is a complete and balanced diet for felids, similar to the diets consumed by wild, free-ranging cats. Although an excellent diet for felids, it is usually expensive and cost-prohibitive to provide sufficient quantities of whole prey for large felids as a daily diet.
FAT
The most concentrated source of energy in the diet is fat, which also gives palatability to foods. Fat provides essential fatty acids and is a carrier of fat-soluble vitamins. The essential fatty acids (linoleic, alpha-linolenic, and arachidonic acids) are involved in many aspects of health, including skin and coat condition, kidney function, and reproduction. Another unusual characteristic of the felid is that essential fatty acid requirements cannot be met solely from linoleic or linolenic acids, as occurs in most mammals studied. In addition, cats require a long-chain fatty acid, arachidonic acid, which is available only from animal sources.22 This requirement appears to stem from low activity of hepatic desaturase enzymes required to convert linoleic to arachidonic acid.
CAROTENOIDS AND VITAMINS
The liver is the major vitamin A storage organ for those species that have been studied. All whole-prey diets (including viscera) analyzed to date would appear to exceed the dietary requirements for cats (∼3333 IU/kg dry matter) (see Tables 33-1 and 33-2)19 without a need for further supplementation. In contrast, all-meat or chicken-neck diets are known to be deficient in essential vitamins, including vitamins A, D, and E.29 Vitamin A deficiencies have reproductive consequences in female felids, primarily pregnancy loss and small litter size.24 Although comparative data are lacking for male felids, vitamins A and E have a pronounced effect on spermatogenesis in other mammals.16,17 A 2-month deficiency of vitamin A was reported to cause endocrine changes and complete aspermia in rats.13,14 Vitamin E deficiency also influences spermatogenic development in the boar17 and causes incomplete spermatogenesis and epididymal dysfunction in the rat.4
Levels of vitamin A, D, and E in various feline diets are listed in Table 33-2.
MINERALS: CALCIUM AND PHOSPHORUS
The cat’s requirements for other nutrients, such as calcium and phosphorus, appear to be similar to those in other mammals. As a food source, a whole-prey vertebrate carcass generally is similar in nutrient composition across species (e.g., rat, mouse, chick, rabbit) and provides adequate amounts of calcium (Ca) and phosphorus (P) and in a satisfactory Ca/P ratio (∼1.5:1). In contrast, muscle meat is different in nutrient composition from whole prey.2 Muscle meat is a good source of protein (see Table 33-2), but it is extremely low in calcium, resulting in an inverse Ca/P ratio Table 33-3.1 In felids, dietary calcium deficiency causes resorption of bone mineral, greatly reduced bone density, and ultimately metabolic bone disease.28 Over time, bone demineralization results in fibrous osteodystrophy and nutritional secondary hyperparathyroidism.25,28
Table 33-3 Calcium (Ca) and Phosphorus (P) in Vertebrate Carcasses, Manufactured Diets, and Muscle Meats Compared with the Estimated Nutrient Requirements (Dry Matter Basis) for the Domestic Cat
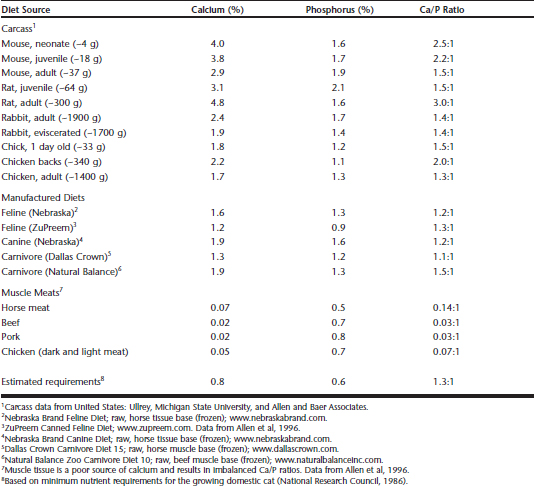
The calcium and phosphorus levels of various whole-prey carcasses and several common commercial diets are listed in Table 33-3. For comparison, muscle meats also are listed to illustrate the calcium deficiency and imbalance in calcium and phosphorus.
IMBALANCED DIETS AND FERTILITY IN FELIDS
Latin American Cats
Data from a reproductive survey of felids in Latin America (Mexico, Central America, South America) demonstrate the importance of diet on reproductive health and breeding programs.27 Reproductive evaluations were conducted on 185 captive, adult male felids representing eight endemic Latin American felid species, including the ocelot (Leopardus pardalis), margay (Leopardus wiedii), Geoffroy’s cat (Oncifelis geoffroyi), tigrina (Leopardus tigrinus), pampas cat (Oncifelis colocolo), jaguarundi (Herpailurus yaguarondi), jaguar (Panthera onca), and puma, that were maintained under a variety of dietary and other management conditions in 44 zoos or private facilities in 12 Latin American countries. Of the 185 males in the survey, 172 (93%) were wild-born. The remaining 13 individuals were captive-born from wild-born parents. Almost all the small cats (126/129, 98%) were wild-born compared with the larger cats (46/56, 82%).
Of the 185 male cats assessed, the level of successful captive breeding was low, with only 37 males (20%) classified as proven breeders (produced at least one offspring).27 The majority of these proven breeders were jaguars, pumas, and ocelots. Reproductive evaluations of these 185 males revealed that 131 males (71%) had sperm in their ejaculates; however, the mean number of sperm/ejaculate for each species was low compared with counterpart values measured in U.S. institutions. More than half of all ejaculates contained less than 1 million total sperm, which is an extraordinarily low number for felids.11,30 Except for the pampas cat, aspermic individuals were observed in all species. Only 87 males (47%) had at least 1 × 106 total sperm/ejaculate, and only 53 males (29%) had 10 × 106 or greater total sperm/ejaculate.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


