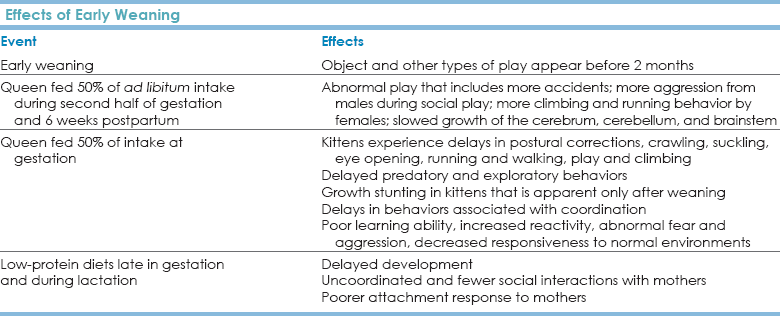
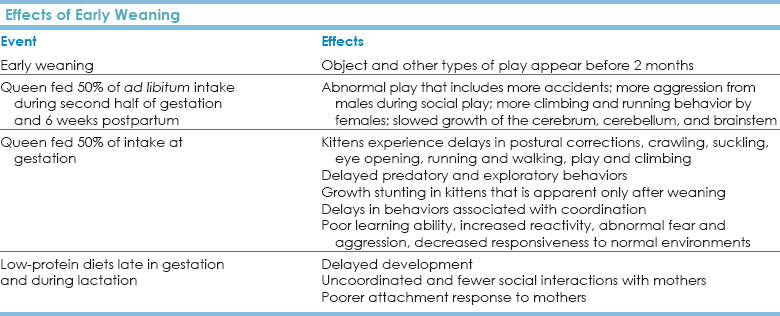
Chapter 8 Evolutionary History and the Cat’s Relationship with People If the story of dogs is the story of collaborative work with humans, the story of cats is one of disease and theft-of-food control. The domestic cat, Felis catus, is derived from Felis libyca, the African wildcat that is said to “tame” when taken into a household. Genomic cladistic analysis has grouped together a cluster of small cats—F. catus with Felis silvestris (the European wildcat), F. libyca (the African wildcat), Felis bieti (the Chinese desert cat), Felis margarita (desert cat), Felis nigripes (the black-footed cat), and Felis chaus (the jungle cat) (Johnson et al., 2006)—but Felis silvestris lybica wildcats from Israel, the United Arab Emirates, and Saudi Arabia are alone among those in the wildcat lineage because they are genetically indistinguishable from domestic cats (Driscoll et al., 2009). This means that domestic cats must have arisen from a single location, the Middle East. None of the cats we now recognize as “domestic” are derived from wildcats in any other locations. This juxtaposition of cats, rodents, and humans likely allowed feline “domestication” about 10,000 years ago, which is 5000 to 25,000 years later than for dogs, using conservative estimates of domestic dog development. The cohabitation of these small, tamable cats and humans occurred at a time when agriculture was beginning to become established in the Middle East. The establishment of agriculture may have been the key that united cats and humans, and both genetic and archeological data support this time period as the one when cats and humans began to live together in an exceptional form of “domestication” (Driscoll et al., 2009). • Feline predatory behavior and most of the other attendant social behaviors that cats exhibit in human households have been only slightly changed in their structure and behavioral elements since the cat was “domesticated.” • Because cats have had little direct influence on human economics (e.g., compared with that of hunting or herding dogs), there has been little intervention in their behavior by humans in the past hundred years (Young, 1985). • Lack of selection for specific suites of behaviors (“behavioral domestication”) also may be reflected in the range of body sizes for present-day cats. In the wild, the range of body sizes of wild cat species greatly exceeds the range of size of wild dog species, and the range of body sizes of domesticated dogs far surpasses that reported for wild canids. In contrast, the size range of domestic cats is relatively small and well within the range of body sizes reported for their wild counterparts. Even large domestic cats are relatively small compared with their wild relatives. • The lack of human intervention is supported by coat color patterns. One of the most common feline coat colors, the tabby, has radiated worldwide, yet this color is extremely similar to that of the European wildcat and Norwegian forest cat. • Dogs, with whom we evolutionarily first formed true collaborative partnerships based on work • Stock, animals such as sheep, goats, cows, chickens, and horses, who work for—not with—and/or serve us in the most utilitarian of contexts (i.e., providing food, fiber, compelled labor, et cetera) These conditions ensured that artificial selection would allow humans to alter physique, behaviors, and physiologies in ways that helped or were desired by humans and that the resultant derivative changes, as much as any of the conditions enumerated by Galton, are responsible for the condition of the domesticated species that we see today. For stock, we have altered looks, physiology, and some aspects of behavior, such as flight distance and tractability. For dogs, where we may well have a true co-evolutionary relationship, as part of selecting dogs for specific tasks and working with them in those tasks, we have facilitated neuromolecular changes that mirror those of humans (Saetre et al., 2004). This process of choosing desirable behaviors and physiologies and altering them in ways favorable to humans is at the core of the process of “domestication,” and it has not been traditionally involved in our relationship with cats until the most recent few hundred years. The process of domesticating cats is actively ongoing and can be seen in the breeds developed in the past 50 years and in changes in coat length and texture, ear shape, social preferences, and physical and vocal behaviors. The misunderstanding of cats is a serious humane concern. Cats continue to rise in popularity as pets worldwide. Cats are now the most popular pet in the world, with a third of U.S. households having at least 1 cat and 600 million cats living with humans worldwide (Driscoll et al., 2009). Domestic Cat Social Grouping and Reproduction The focus of feline social groups is invariably the female and her kittens. Most studies have concentrated on matrilineal relationships as the focus of the social grouping. Similar to lion prides, domestic cat groups are often composed of related females and their juvenile offspring. It has been postulated that this social pattern is driven by the reproductive pattern of female cats. See Table 8-1 for a summary of reproductive hallmarks. TABLE 8-1 Hallmarks of Feline Reproductive Biology • Domestic cats reach puberty at about 10 months of age, although females born in the late spring may not cycle until the following year. • Peaks in sexual activity occur from mid-January through March and from May to June in the Northern Hemisphere. • Domestic cats are seasonally polyestrous and generally experience two cycles per year if not bred. Domestic cats can cycle every 3 weeks for several months—a fact that may help the most reluctant client to consider neutering the cat. Estrus in cats lasts 9 to 10 days in the absence of copulation but only 4 days if copulation occurs because cats are induced ovulators. • Gestation in female domestic cats is approximately 63 days. This is longer, by 3 to 7 days, than the gestation period for F. sylvestris libyca (Hemmer, 1979; Schmidt et al., 1983). Litter sizes vary from 1 to 10 kittens with a mean of 4.5 kittens per litter. Most domestic cats allowed to do so have two litters per year (Liberg and Sandell, 1988), so one cat may produce an average of nine kittens a year. It is easy to understand how cities or neighborhoods with feral or semi-feral stray cat populations can be overwhelmed within a few years if the cats are left intact and have some access to shelter and food. • In one cohort study on the effect of trap-neuter-release (TNR) programs designed to decrease the number of unowned/feral cats for humane, public health, and conservation reasons, the mean number of kittens per litter was three (range, one to six). Fetus counts indicated a median litter size of 4, with a range of 1 to 10 kittens as reported (Liberg and Sandell, 1988). Before 6 months of age, 127 of 169 (75%) kittens born to the unneutered cohort died or disappeared, with trauma as the most common cause of death (Nutter et al., 2004). • Cats usually have eight nipples. Only three pairs of these nipples may produce sufficient milk to provide nourishment for a kitten, and the back nipples are preferred by kittens (Rosenblatt, 1976). Teat preference is established by 1 to 3 days, when present, and 80% of all kittens develop a teat preference (Rosenblatt et al., 1961). Kittens identify their mother and their preferred nipple using innate and learned olfactory cues (Raihani et al., 2009). The effects of such patterns on early nutrition and neurodevelopment have not been investigated, but for cats who may be compromised, this pattern may lead to some kittens being under-nourished, a pattern that contributes to increased reactivity and later problematic behaviors (Carola and Gross, 2010; Green et al., 2011). • Females form stable matriarchal groups and may join in communal nests and engage in shared care and nursing (Macdonald and Apps, 1978; Macdonald et al., 1987). The queen moves the kittens if (1) the nest is fouled by feces, (2) undigested prey remains as the kittens get older, or (3) the site is disturbed by a strange male cat (Leyhausen, 1979). • Queens stimulate kittens to eliminate by the anogenital reflex until 23 to 39 days, and kittens can usually eliminate voluntarily by 3 weeks. This developmental change may be associated with the queen’s mobile behaviors. Cats move kittens most frequently between 25 and 35 days (Schneirla et al., 1963). • A familiar male may be such an integral part of the social group that he will guard against disturbances of foreign males (Macdonald et al., 1987). Communal nests may function in part to repel intruder males; however, they may also function to ensure that all kittens receive nourishment, should the number of nipples or the production of the milk be insufficient to support large litters. In a large farm study, males would provide care and succor for the kittens, if these males were residents within the larger social group (Macdonald et al., 1987). • The presence of several reproductive males may facilitate communal care, including communal care provided by males (Kerby and Macdonald, 1988; Turner and Mertens, 1986). In such cases, no “dominant” male monopolizes all the matings (Natoli and de Vito, 1988). If males can neither monopolize all matings nor guarantee paternity, it might be advantageous for them to contribute to communal care. Many studies have shown that they are close to and will collaborate with females who are sisters and mothers (and possibly grandmothers) (Curtis et al., 2003). Males seldom provide lactating females with food or bring food directly to the kittens (Liberg, 1980), but they may care for the kittens, washing, guarding, and “nursing” them in non-nutritive relationships. It is now established that cats have “preferred associates” (Curtis et al., 2003), but these relationships go beyond the purported genetic ones. • Females may further add to the stability of such social systems through their behavior during proestrus. During proestrus, females are attracted to males but are not receptive to their courtship behavior and reject attempts at copulation. • It has been postulated that this is a device to enhance male-male competition (Bradshaw, 1992). • This proestrus behavior also provides females with the opportunity to assess the staying power and abilities of males. In colonies that are sufficiently large, a proestrus that allows the females to evaluate males that are attracted to them would be one mechanism by which they could identify resident males and males with skills that may contribute to the best outcomes for the young. • Finally, if the females are attractive to a number of males and all those males are part of the colony, that behavioral interaction could help facilitate communal care. In such cases, there may be an element of uncertainty of paternity, but copulation could then act to solidify social relationships where individuals “within” the group are treated differently than individuals considered “outside” the group. There are no data on whether animals known to the group are less likely to practice infanticide or more likely to provide communal care; there are even fewer data on whether the extent of a genetic or social relationship makes a difference for either of these factors. It has been postulated that males within colonies could be related. If this were the case, infanticide would not be an appropriate genetic strategy and would likely not enhance any single male cat’s genetic contribution because insemination by a brother would also be genetically advantageous. • Relatedness has become a fascinating issue because ovulation is induced and estrus for any queen lasts only about 4 to 5 days (Natoli and de Vito, 1988, 1991; Paape et al., 1975; Schmidt et al., 1983). • Liberg (1983) noted that at the peak of fertilization the queen was monopolized by the “dominant” male in the group. Females can thwart this guarding by allowing younger, lighter males to mate before mating with the older, heavier male, resulting in some mixed paternity litters, and colonies where almost half the kittens are not fathered by the putative “dominant” male (Yamane, 1998). • A 24-hour capacitance period is necessary for cat sperm, and the female ovulates 24 hours after copulation (Hamner et al., 1970), so a solitary copulation may not be sufficient to induce females to ovulate and to ensure that sperm capacitance occurs at the time of ovulation. • This physiological pattern could argue strongly for either the guarding effect noted by Liberg (1983) or the effect hinted at by Bradshaw (1992) and found by Yamane (1998) in which multiple males might possibly contribute to insemination and, indirectly, to the stability of the social group in groups where males provide a portion of the communal care for the young. It is conceivable that both modes function depending on the demography of the group and the resources available to it. Social structure is influenced by both the age and the sex structure of the group and will interact with the resource, predation, and physical and biophysical environments to affect behaviors. • The issue of infanticide may be a concern in free-ranging groups and in breeding catteries. In domestic cats, weaning normally starts at approximately day 30 and continues through day 60 of the kitten’s life. If kittens die, regardless of the cause of death, the queen enters estrus approximately 15 days later (Liberg and Sandell, 1988). Under normal circumstances, cats might have two litters a year; if there is a litter death, the inter-birth interval can be only 133 days. Infanticide in domestic cats has been rarely observed directly (for exceptions see Dards [1983] and Liberg [1983]). An evolutionary argument can be made for the occurrence of infanticide in unstable groups or in groups that experience a cataclysmic change in the social structure. Females are induced ovulators, and the basis of social communal relationships may be influenced by the relatedness of the colony (Packer et al., 1991). There are no hard data to support this view of infanticide, and it would be inappropriate to assume that all of the social relationships within feline groups are driven by direct genetic relationships. Kittens in wild groupings often stay with their mother until at least 6 months or 1 year of age and then disperse. During this intermediate period, should they stay with the mother or with the mother’s social group, the kittens often form independent social associations (Leyhausen, 1979; Macdonald et al., 1987). After approximately 10 to 14 months of close association with the mother and her social coterie, the kittens experience what has been described as a “social” weaning from the group (Wolski, 1982). For cats, we lack many of the detailed ontogenetic data available for dogs, but the landmarks that are well established are included here and summarized in Table 8-2. TABLE 8-2 Developmental Hallmarks and Periods, including Sensitive Periods and Associated Neurodevelopmental and Behavioral Landmarks* *Note: These periods indicate when cats are first most receptive to the noted stimuli. There is no implication that cats ever stop learning from their experience. See text for references. • The tactile sense is present early in gestation. • Kittens are born unable to see but locomote freely. • Kittens 2 to 4 weeks old exhibit plasticity physiologically and behaviorally. Exposure to low temperature can influence the rate of temperature regulation development at 2 weeks of age, but this effect is lost by 4 weeks (Jensen et al., 1980). This observation reinforces the extent to which early ontogeny is important in cats. • Eye opening, between 10 and 14 days, is affected both by the environment and by genetics. Kittens of young mothers appear to open their eyes earlier than kittens of older mothers. Female kittens tend to open their eyes slightly earlier than males, but there also appears to be a strong paternal genetic effect on timing of eye opening (Bateson and Turner, 1988; Turner et al., 1986). This is probably not independent of the paternal effect on “boldness” (McCune, 1992). • Electroencephalogram (EEG) activity in cats has periods of quiescence at 2 weeks, but EEG activity is continuous by 4 weeks. A peak in EEG activity occurs between 6 and 8 weeks, suggesting a neurodevelopmental shift, but amplitude and absolute power of EEGs do not reach approximate adult values until 20 to 24 weeks of age (Lewis et al., 2011). • Kittens can recognize their mothers by sight and smell by the end of the third week of age (Martin and Bateson, 1988). • By about 6 weeks of age, kittens demonstrate an adult-like response to visual and olfactory social stimuli, including silhouettes of adult cats and the scent of adult cat urine (Kolb and Nonneman, 1975). At about this time, the gape (flehmen) response to cat urine appears; this becomes fully expressed by 7 weeks of age. It has been postulated that the gape response, in its full adult form, is an indication of the maturation of the vomeronasal organ. • The classic adult threat posture of tail down, arched back, and erect ears is often seen concomitant with the development of the gape/flehmen response and appears at about the same time (Kolb and Nonneman, 1975). • Kittens can learn from firsthand, trial-and-error experience and by watching other cats. They are quicker at observational learning if they are watching their mother (Chesler, 1969; John et al., 1968), suggesting that there may be a role for very elegant, complex, and detailed signaling behaviors that have not been studied. • The role of early social exposure in learning is also suggested by experimental results derived from maze problem-solving tests (Hebb-Williams closed field test). Alley cats achieve much higher scores than housecats, but they also make more errors (Pollard et al., 1971). • Cats isolated from other cats from birth until 7 months of age are slow to accept introduced cats (Kolb and Nonneman, 1975; Konrad and Bagshaw, 1970). • Kittens who are isolated from other kittens until late in their first year of life display exaggerated autonomic responses characterized by galvanic skin responses and disruption of regular sleep rhythms (Wenzel, 1959). • Neonatal isolation leads to changes in normal pain response in dogs (Melzack and Scott, 1957), but it is correlated with increased aggression in cats and rats (Guyot et al., 1983). • Kittens artificially separated from their mothers at about 2 weeks of age become fearful and aggressive toward both other cats and people, demonstrate random locomotion, and learn poorly (Bacon, 1973; Seitz, 1959). • Singleton kittens are quicker to emerge from nest boxes, between 3 and 7 weeks (Mendl, 1988), and appear to show little distress when left alone. • Two kittens, when left by their mother, appear all right while together, but if the kittens are separated and then left by their mother, they appear more distressed than a singleton kitten that is left alone (Mendl, 1988). Most variation in individual behavior is probably not attributable to litter composition (Deag et al., 1988). • By the time the kittens are about 4 weeks of age, the queen starts to bring them solid food, representing the beginning of the weaning phase (Ewer, 1961; Kovach and Kling, 1967; Martin and Bateson, 1988). This phase is completed by 7 weeks of age. • Kittens who are nursed on a breeder nipple nurse normally but will not be permitted to nurse on a lactating queen because they give inappropriate social responses to her (Rosenblatt et al., 1961). These kittens can form social attachments to other kittens, but they do so slowly. The extent to which these slow social attachments and inappropriate social responses to lactating queens may affect their own ability to raise kittens has not been investigated. Such scenarios might be an important factor in the abandoned, feral cat population in which a decreased plane of nutrition and early abandonment may be common. • Kittens who are weaned early also develop predatory behavior far earlier than do normally weaned kittens and are more likely to be mouse killers (Tan and Counsilman, 1985; Warren and Levy, 1979). Late weaning of kittens appears to delay predatory behavior and decreases the propensity to kill mice. This association may be indicative of nonspecific learning in response to the most common prey (mice), but the implications of this are important for people who are going to hand-rear cats. People who hand-raise early-weaned kittens might experience problems with earlier predatory behavior. • By about 5 to 6 weeks of age, kittens are totally independent in their ability both to eliminate and to start to find substrates resembling the ancestral substrate for elimination. • For the first 3 to 5 weeks, the queen stimulates the kittens to eliminate and cleans up after them. • By 3 to 5 or 6 weeks of age, kittens begin to seek open, well-drained substrates and use them for both urination and defecation (Fox, 1975). • Covering of urine or feces may not occur—the ancestral condition is to spray urine a large percentage of the time, and feces may not be covered in many arid environments. Feces are often used as signposts for territorial delineation (Wemmer and Scow, 1977), but it may also be possible to make a case for disease control when feces are left exposed. When feces are exposed, disease transmission is decreased, and parasites are killed in arid environments. • If the cats are going to cover their urine and feces, they appear to start to do so about the same time they are able to respond fully to an olfactory challenge, at approximately 7 weeks of age. No quantitative data exist for age-specific elimination behaviors in cats. • As late as 13 to 18 months of age, young males may only squat to urinate (Wolski, 1982), suggesting a strong social component for spraying. It is rare for free-ranging males to show urine marking in paternal areas. Housecats who are not interacting in these social environments usually stop solely squatting at about 8 to 10 months of age. • The most significant side effect of early weaning might be on play behavior. Under normal conditions, object play increases by the second month (Bateson, 1978; Dumas, 1992), but under early-weaning conditions generated by gradual separation, the administration of bromocriptine (a dopamine antagonist that stops lactation) (Bateson and Young, 1981), or decrease in the maternal food supply, certain types of play show an early increase (Bateson and Young, 1981; Martin, 1984; Martin and Bateson, 1985). These changes may be adaptive responses to forced independence. • Queens fed 50% of their ad libitum intake during the second half of gestation and for the first 6 weeks postpartum produce kittens with abnormal play behavior. Afflicted kittens have more accidents during play, the males demonstrate increased aggression during social play, and the females demonstrate less climbing and more running behavior; also, the cerebrum, cerebellum, and brainstem do not appear to grow at the same rate as the rest of the brain. The brain and body appear to achieve normal size once rehabilitated with food, but the extent to which there might be long-lasting effects on attachment has not been fully explored (Smith and Jansen, 1977a, 1977b, 1977c). • When maternal food intake at gestation is 50% of normal, this produces kittens with delays in postural corrections, crawling, suckling, eye opening, running and walking, and play and climbing (Simonson, 1979). These kittens also have delayed predatory and exploratory behaviors and experience the growth stunting that becomes apparent only after weaning. The greatest behavioral delays appear in behaviors that regulate coordination, and these kittens have poor learning ability, increased reactivity, abnormal fear and aggression, and a decreased responsiveness to normal environments. • Low-protein diets late in gestation and during lactation correlate with delayed development; kittens that experienced these restricted diets are uncoordinated and exhibit fewer social interactions with their mothers and poorer attachment responses to their mothers (Gallo et al., 1980, 1984). • Social relationships that provide the context for future communal suckling develop within the first 2 months after birth (Macdonald and Apps, 1978). These relationships are adversely affected by these nutritional situations. • Cats who are abandoned and may become feral or cats born to mothers that are abandoned and possibly feral develop abnormal social behaviors. The combination of poor learning, increased reactivity, out-of-context and more intense than normal reaction in any foreign circumstance, abnormal fear accompanied by aggression, and decreased ability to respond in normal situations may render these animals poor candidates for both rehabilitation and for pets. “Good Samaritans” should be aware of these associations and have realistic expectations. • Well-nourished male and female kittens are similar in mass and behavior until about 8 weeks of age. Thereafter, males are larger than females (Liberg, 1983). Males take 3 years to reach their adult weight, whereas females take only 2 years. This occurrence is associated with the period during which cats mature socially. • Under normal conditions, feline tactile sensitivity is fully developed by the 24th day of gestation, and the vestibular righting reflex is developed by the 54th day of gestation. It is unknown how early in gestation the effects of dietary restriction are manifest. These findings are in line with findings from research showing that early and in utero deprivation cause long-lasting detrimental effects on behavior, promoting hyper-reactivity and anxiety (Carola and Gross, 2010; Green et al., 2011). We do not know whether there are epigenetic effects that shape behavior transgenerationally as has been found in other species (Radtke et al., 2011), but there is no reason why cats should be exempt. Such risks may be most profound for feral and/or homeless cats. The effects of weaning on behavior are summarized in Table 8-3. The play behavior of kittens has been extensively studied, in part because of the potential association between play and predatory behavior in cats. The rich, imaginative type of play in which kittens engage is one of the appeals of having a kitten. Although play changes with age (Table 8-4), cats do not stop playing as they age if they have someone with whom to play. The character of the play alters, but cats can continue to be playful given the appropriate stimulation. TABLE 8-4 Developmental Hallmarks in Play and Predatory Behavior • Sex differences in social play appear by weeks 12 to 16 but are not present between weeks 4 and 12 (Barrett and Bateson, 1978). • Females who play with males become more male-like in their play behavior than females who play with females (Caro, 1981a, 1981b). Whether this factor affects later aggressive play behavior for females raised with male siblings only has not been investigated. • Normal social play starts at approximately 3 to 4 weeks of age when the kitten can ambulate. It is honed when eye-paw coordination is developed at approximately 7 to 8 weeks of age (Barrett and Bateson, 1978; Martin and Bateson, 1985). • Complex motor activity is fully functional by 10 to 11 weeks (Villablanca and Olmstead, 1979). • Social play begins to decline at about 12 to 14 weeks of age (Caro, 1981a; West, 1974, 1979). • Social play patterns become more associated with predatory behavior and social fighting by the third month, possibly concomitant with a change in systems that control motor behavior (Caro, 1980b, 1981b; Pellis et al., 1988; West, 1979). • Many motor patterns in play resemble motor patterns for hunting (Caro, 1979, 1980a, 1980c), but there is no definitive link with play as “practice” behavior for predatory skills and the development of those skills later in life except to the extent that we all learn from experience in any social situation in which we are placed (Martin, 1984). • Cats deprived of play still develop predatory behavior (Baerends-van Roon and Baerends, 1979). • Cats who did not or could not play with small objects as kittens appear to be no different than other cats when examined with regard to predatory skills at 6 months of age (Caro, 1980b), but cats are still more likely to kill the types of prey that they had known since kittenhood (Caro, 1980a). • The predisposition to respond defensively toward large and difficult prey develops some time during the second month (Adamec et al., 1983). • Free-ranging domestic cats start to bring their kittens live prey about 4 weeks of age, coincident with the beginning of the weaning. • Kittens can start to kill mice on their own at approximately 5 weeks of age (Baerends-van Roon and Baerends, 1979). • It is not inevitable that cats will hunt, especially if they are not weaned early and if they are not taught to do so by their mothers. The presentation of palatable food starting at kittenhood may inhibit hunting, although the behaviors of killing and eating are separately, centrally controlled (Adamec, 1975b, 1976a, 1976b). • Although individually controlled, there is some interaction between killing and eating, which is substantiated by interactions between areas of the lateral and ventromedial hypothalamus. The ability to inhibit hunting through the presentation of palatable foods early in life and throughout life may be important for clients who do not wish their cats to hunt. • The tendency to kill when hunting or to exhibit hunting behavior increases with hunger (Biben, 1979) and decreases when prey is more difficult to capture. • Population control at the community level plays a role in hunting behaviors. Females with kittens to feed tend to be far more adept hunters than females who do not have kittens (Turner and Meister, 1988). It has been postulated that this facilitation is a dopaminergic/prolactin response. • Weanling kittens eat food preferred by their mother even though that food may not be a common food in cat diets (Wyrwicka, 1978). • Kittens appear to be able to learn to kill a rat by watching another cat do so (Kuo, 1930). They also learn a preference at this stage and restrict their preference to the strain of rat with which they are familiar. • Competition within litters may also hone some predatory skills (Caro, 1980a, 1980b, 1980c), and some cats appear to be able to “catch up” to more skilled littermates during ontogeny (Caro, 1980b). • Cats can learn about a variety of tasks through observation and may learn more quickly if it is their mother they are observing. This finding has profound and helpful applications for clients adopting kittens who may also be able to adopt the mother. • The tendency for cats to retrieve their young in response to high-pitched vocalization peaks at 1 week after parturition (Schneirla et al., 1963). Early in life (during the first 3 weeks), kittens use ultrasonic calls as they explore their nest (cited in Deag et al., 1988). These calls may function in helping their caretakers locate them and keep them together. • Eight kitten play postures have been identified (West, 1974): • Behaviors, some of which are associated with later predatory behavior, that are most successful in eliciting play from another kitten are the pounce (39% of all play-eliciting behaviors), the belly-up display (14% of all play-eliciting behaviors), and the stand-up (16% of all play-eliciting behaviors). These behaviors are 90% effective in obtaining a response from another kitten between 6 and 12 weeks of age. • As the cat matures, other behaviors become important in the play elicitation communication repertoire. The vertical stance elicits play behavior only 8% of the time at 6 weeks of age, but by 12 weeks of age it elicits play response from another littermate 24% of the time. In contrast, the side-step posture elicits play 20% of the time at 6 weeks of age but only 3% of the time at 12 weeks of age (West, 1979). It is important to emphasize these normal changes in communicatory and play behavior to clients because potential problems in communication could arise when people play too roughly with their kittens. Clients need to know appropriate, age-specific play behaviors. • In his study of singleton litters, Mendl (1988) found that singleton kittens did not engage more frequently in self-play or object play than kittens in litters of two. This finding suggests that social and object play should be separately driven. • Solitary kittens directed all of their play toward the mother; given that the mother is more likely to be absent from the nest than a sibling would, singleton kittens might experience less social play. • Between 2 and 4 weeks of age, the particular closeness experienced by kittens to other kittens has a calming effect (Rosenblatt et al., 1962). • Physical contact with the queen, particularly nuzzling of the face, has a calming effect on kittens (Beaver, 1992). These observations have implications for early weaning. • Kittens are weaned early if the queen cannot provide enough food; this results in stimulation of the early development of both object and social play (Bateson et al., 1990). • In the case of early weaning, the siblings and the mother are still present; their presence could have a significant effect on the extent to which rough play might be modulated. In the usual, non-experimental situation involving early weaning, the mother is no longer present. No studies have been done on whether these two types of early weaning have different effects on inappropriate play behavior or the early development of predatory aggression in cats. It is likely that early weaning that also precludes the social interaction of an older, experienced cat could foster more inappropriate play behavior and play aggression. • Feral cats often abandon kittens by about 4 months of age. At this time, the kittens increase their environmental exploration. • Barrett and Bateson (1978) noted that males engage in twice the object play as females by 7 weeks of age, and by 19 weeks of age these free-ranging males are demonstrating sexual behavior. • Female kittens usually do not demonstrate sexual behavior until 23 weeks of age. Litters composed solely of females appear to be less aggressive in their play than all-male litters; this difference is recognizable by weeks 12 to 16. • West (1979) postulated that the function of play, rather than teaching cats to be better predators later in life, might be to keep litters together when the litter is vulnerable; this would enhance their ability to develop good social relationships that might serve to redress vulnerability later in life. No studies have directly addressed this issue, but the work of Macdonald et al. (1987) suggests that this might be so. • Odor is sufficiently important for feline feeding behavior that the odor of a palatable food (cooked rabbit) can initiate feeding in the absence of any change in food offered (Robinson, 1992a). • Moisture determines meal size and eating speed (Robinson, 1992a). If a cat’s diet is diluted with water, the cat compensates by eating more (Castonguay, 1981; Mugford, 1977). This effect is not found if inert substances such as kaolin or cellulose are added to the diet (Hirsch et al., 1978; Kanarek, 1975)—hence the basis for low-calorie cat foods. • Wet food is initially consumed quickly, followed by a subsequent decrease in feeding rate. Dry, calorically dense food is consumed at a slower, more consistent rate. It is unclear whether the costs of handling affect this comparison. • The preferred food temperature is 35° C—this may be the temperature that most effectively releases volatile fatty acids. • Cats gain and lose mass cyclically (Randall and Lasko, 1968). This pattern may be associated with their annual cycles of corticosteroid, thyroxine, and epinephrine, which peak in the winter (Anderson, 1973; Randall et al., 1975). Regardless, cats appear to regulate the intake of energy, not volume or mass (Robinson, 1992a). For commercial dry foods (360 kcal/100 g), an average meal contains 22.7 to 31.3 kcal. Cats eat an average meal of 35.5 kcal from fresh meat (136 kcal/100 g), 30.2 to 44.8 kcal from canned meat (80 to 90 kcal/100 g), 19.8 to 32.5 kcal from canned meat and cereal (115 kcal/100 g), and 30.1 kcal from semi-moist food (320 kcal/g) (Robinson, 1992a). • One unpleasant food experience can lead to rejection of that food for months (Houpt, 1982; Macdonald et al., 1984, 1985). Cats have a requirement for thiamine (particularly with a carbohydrate-rich diet). The first symptom of thiamine deficiency could be anorexia. A single arginine-deficient meal can lead to ammonium intoxication, causing emesis and lethargy. Such factors are usually redressed by commercial cat foods, but young animals with abnormal eating patterns may not ingest sufficient food to modulate these concerns. A thorough investigation is warranted in the case of any young kitten that does not eat and thrive. In 1937, Lorenz defined a “critical period” with regard to imprinting. Implicit in the definition was a definitive onset and offset. During these critical periods, animals were postulated to be able to learn to respond to certain stimuli, and before and after these periods animals were unable to respond to the stimuli. The concept of a critical period was modified by Bateson (1979). Instead, Bateson defined a “sensitive period” as an age range during which particular events are especially likely to have long-term effects on individual development. A sensitive period may best be defined as the period of time when you are best able to attend to and be affected by the relevant stimuli, which, when missed, puts you at risk for concerns pertaining to those stimuli. This concept is particularly relevant for developing parts of the nervous system (i.e., visual cortex) that rely on stimuli to direct their development (Rauschecker and Marler, 1987). For example, exposure to contours of only one orientation can have long-term effects on the visual system. Whether any period is “critical” versus “sensitive” may be a matter of our scale of measurement. We should be mindful that there is almost always more variation and plasticity in any behavior and its development than is commonly thought. • Animals are not behaviorally or developmentally able to respond to all stimuli when they are born. • They can begin to respond to certain stimuli within certain broad periods. • There is a considerable amount of variability in response to specific stimuli both within and between litters. • Missing the appropriate stimuli (those to which the individual is now capable of responding) during these periods does not guarantee a “poorly socialized” animal; however, the risk of inappropriate contextual responses increases with increased deprivation. • Animals should be exposed to all relevant social stimuli early and in a non-traumatic manner. When the individual animal is developmentally ready to learn from the stimulus, it will do so. • No harm results from the presence of any stimulus (i.e., other cats, humans) before the time that the animal is best able to attend to it, if no undue trauma or fear is involved. • Animals experiencing all of the appropriate “socialization” may still have behavioral problems. • Although sensitive periods have been less emphasized in the importance of the development of good pet cat behavior than have sensitive periods in the importance of good pet dog behavior, feline sensitive periods may be shorter, more discreet, and more frequently legitimately implicated in the development of behavioral problems such as play aggression, inappropriate play behavior, and fear aggression. • Karsh (1983a, 1983b, 1984) provided baseline data about “sensitive” periods and defined the specific behavioral changes that can occur within the above-outlined time frames for kittens reared in a laboratory situation. • Kittens that were handled by people for only 15 minutes a day from birth through 12 to 14 weeks of age spent more time exploring the person and giving head rubs and would leave and return several times. • Kittens in home-reared litters that were held 1 to 2 hours a day, if brought to the laboratory, would go directly to people and climb onto their lap, purr, and go to sleep. These behaviors were not seen in the laboratory kittens, although laboratory kittens were not fearful of people. The home-reared kittens were handled four to eight times longer than the laboratory kittens, but they were also exposed to a more varied and unpredictable environment than the laboratory kittens. • It would be inappropriate to over-interpret these results, but it is probably fair to say that the earlier the kittens are handled and the more they are handled, the more friendly they are likely to be.
Normal Feline Behavior and Ontogeny
Neurological and Social Development, Signaling, and Normal Feline Behaviors
Overview of Normal Cat Behavior
Behavioral Ontogeny in Cats
Event
Time/Age
Males reach puberty
10 months
Females reach puberty
10 months—if born in late spring may delay cycle until following year
Minimum cycling frequency
3 weeks
Length of estrus with copulation
4 days
Length of estrus without copulation
9-10 days
Gestation
63 days—3-7 days longer than F. silvestris libyca
Capacitance period for sperm
24 hours
Time when ovulation occurs after copulation
24 hours
Re-entry into estrus if kittens die
15 days after their death
Weaning Period
Early Neurobehavioral Development
Developmental Hallmark or Ability
Period/Age
Tactile sensitivity fully developed
Day 24 of gestation
Vestibular righting reflex developed
Day 54 of gestation
Teat preference established
1-3 days
Purring begins
2 days
Eyes open
10-14 days
High-pitched calls by kittens to summon queen peak
1 week
Age at which separation from mother leads to fearful and aggressive behavior to cats and humans
2 weeks
Age at which exposure to low temperature influences the rate of temperature regulation development
2 weeks
Periods of quiescence found in EEG activity
2 weeks
Age at which ultrasonic calls are given by kittens when exploring the nest
Through 3 weeks
Age at which exposure to low temperature ceases to influence the rate of temperature regulation development
4 weeks
Closeness of other kittens has a calming effect
2-4 weeks
Kittens able to recognize mothers by sight and smell
End of 3 weeks
Queen begins to teach predatory behavior
3 weeks
Queen stimulates elimination via anogenital reflex
23-29 days
Kittens eliminate voluntarily
3 weeks
Age at which singleton kittens emerge from nest box
3-7 weeks
Queen starts to bring kittens solid food
4 weeks
Free-ranging cats bring kittens live prey
By ~4 weeks
Normal social play behavior starts
3-4 weeks
Age at which if kittens are exposed to another species (e.g., Chihuahua dog), they will show no fear at 12 weeks
4 weeks
Age through which kittens cannot retract their claws
4 weeks
Age at which kittens will use scratching material if it is provided
5 weeks
Kittens can kill mice on their own
~5 weeks
Kittens moved by queen most frequently
25-35 days
Early period for social play
2-5 weeks
Defensive response toward large and difficult prey develops
During month 2
Kittens independent in their ability to eliminate and find substrates suitable for elimination (the appropriate materials should be provided)
5-6 weeks
Adult-like response to visual and olfactory stimuli, including to silhouettes of adult cats and adult cat urine
~6 weeks
Middle period for social play
5-7 weeks
Gape/flehmen response appears
~6 weeks
Gape/flehmen response fully developed
7 weeks
Age by which if kittens are handled regularly, they will approach unfamiliar objects rapidly and spend more time with them at 4-7 months
Birth to 45 days
Age at which if kittens are handled by numerous people (five people in the study), they will show less fear and more interest in people later
5.5-9.5 weeks
Peak in EEG activity
6-8 weeks
Kittens can respond to an olfactory challenge
7 weeks
Kittens begin to cover their urine or feces if they are going to do so
7 weeks
Queen ceases to bring kittens solid food
7 weeks
Male kittens engage in twice the object play as females
By 7 weeks
Eye-paw coordination honed/object play
7-8 weeks
Late period for social play
7-10 weeks
Kittens completely weaned
60 days
Male and female kittens similar in mass and behavior
Until ~8 weeks
Male kittens become larger than females
Beginning at ~8 weeks
Social relationships that provide the context for future communal suckling
During first 2 months
Object play increases
By 60 days
Complex motor activity fully functional
10-12 weeks
Milk quality changes
12 weeks
No sex differences in social play behavior
4-12 weeks
Pounce, belly-up, and stand-up displays 90% effective in obtaining a play response from another kitten
6-12 weeks
Social play patterns become more associated with predatory behavior and social fighting
By 12 weeks
Handling kittens for only 15 minutes a day produces kittens more solicitous of people
Birth to 12-14 weeks
Post–social play period
14 weeks
Social fighting may start
14 weeks
Sex differences appear in social play behavior
12-16 weeks
Litters composed entirely of females are less aggressive in their play than all-male litters
12-16 weeks
Social play behavior begins to decline
12-14 weeks
Feral cats abandon kittens
As early as 4 months
Free-ranging males demonstrate sexual behavior
19 weeks
Amplitude and absolute power of EEGs decrease to adult levels
20-24 weeks
Free-ranging females demonstrate sexual behavior
23 weeks
Male cats reach adult weight
3 years
Female cats reach adult weight
2 years
Age at which male cats will stop squatting and start spraying unless inhibited from doing so by the social environment
8-10 months
Puberty
10 months
“Social weaning” from maternal group may occur
10-14 months
Effect of Deprivation on Ontogeny
Effects of Litter Size on Ontogeny
Factors Affecting Ontogeny of Feeding Behaviors
Ontogeny of Elimination Behaviors
Ontogeny of Social Behavior and Role for Nutrition
Ontogeny of Play and Predatory Behavior
Age
Hallmark Behaviors
3-4 weeks
Social play begins
4 weeks
Free-ranging queens begin to bring kittens live prey as weaning begins
5 weeks
Kittens can start to kill mice on their own
5-8 weeks
Predisposition to respond defensively toward large and difficult prey develops
6 weeks
Vertical stance elicits play from a littermate 8% of time
7 weeks
Males engage in twice the object play that females do
7-8 weeks
Real honing of social play associated with development of eye-paw coordination
4-12 weeks
No sex differences in social play
12-16 weeks
Sex differences in social play appear
12-16 weeks
Litters comprising only females play differently than all-male litters
10-11 weeks
Complex motor activity fully functional
12-16 weeks
Social play declines
≥12 weeks
Social play patterns change into social fighting and predatory behavior
6 weeks
Side-step posture elicits play 20% of the time
12 weeks
Side-step posture elicits play 3% of the time
6-12 weeks
Pounce, belly-up, and stand-up behaviors are 90% effective in eliciting play from another kitten
12 weeks
Vertical stance elicits play from a littermate 24% of the time
4 months
Age at which feral cats may abandon their kittens
6 months
Age at which cats that did and did not play with small objects as kittens were tested for differences in predatory skills; no difference was found
Factors Affecting Feeding Behaviors
Early Social Development and Age-Specific Effects on Friendliness and Exploratory Behavior
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Normal Feline Behavior and Ontogeny: Neurological and Social Development, Signaling, and Normal Feline Behaviors