Musculoskeletal Surgery
Musculoskeletal Anatomy
Anatomy of the Suspensory Apparatus and Distal Limb
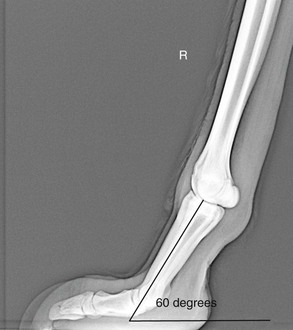
The llama has two digits per limb and, unlike other ungulates, bears weight on both the second and third phalanges. Because of this, both the proximal and distal interphalangeal joints are vertically oriented. The llama does not have a distal sesamoid bone (navicular bone). Each digit has a pair of proximal sesamoid bones, making a total of four per limb. The overall angle of the distal limb is upright, with the normal angle of the first phalanx to the ground being around 60 to 65 degrees (Figure 58-1).1
Individual articulations exist between the medial and lateral first phalanges and the single fused cannon bone of each limb (metacarpal III and IV and metatarsal III and IV). Because of these individual articulations, the fetlock joint has two compartments (medial and lateral), which are separate most of the time.2 The suspensory apparatus supports the fetlock joint and consists primarily of the suspensory ligament (interosseous muscle) and distal sesamoidean ligaments, with additional support from the superficial and deep digital flexor tendons (DDFTs).
Suspensory Ligament
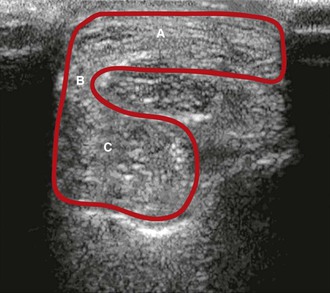
The llama’s suspensory ligament originates from several sites: the second row of carpal bones with attachments to the accessory carpal bone, the caudodistal aspect of the radius, and, most significantly, the palmar aspect of metacarpal III and IV (Table 58-1). The thick palmar fascia connects the suspensory ligament to the superficial digital flexor (SDF) tendon on the lateral aspect of the fused metacarpals at the level of the origin of the suspensory ligament. This can be visualized with ultrasonography (Figure 58-2). The suspensory ligament separates into separate branches proximal to the fetlock joint, with insertions on each of the four sesamoid bones. Additionally, small branches insert on the proximal aspect of the first phalanx. The suspensory ligament is similar in the hindlimb, except that the origin occurs at the proximal plantar aspect of the fused metatarsal III and IV and the plantar aspect of the distal row of tarsal bones.3,4
TABLE 58-1
Anatomic Origin and Insertion Points for Soft Tissue Structures of the Distal Limb in Llamas and alpacas
| Structure | Origin | Insertion |
| Suspensory ligament | Distal row of carpal bones; caudodistal aspect of radius; palmar aspect of fused metacarpus III & IV | Proximal aspect of proximal sesamoid bones; first phalanx |
| Short sesamoidean ligament | Sesamoids | Corresponding first phalanx |
| Cruciate sesamoidean ligament | Sesamoids | Corresponding first phalanx |
| Oblique sesamoidean ligament | Sesamoids | Palmar aspect of first phalanx |
| Interdigital intersesamoidean ligament | Connects two axial sesamoids | |
| Interdigital metacarpophalangosesamoidean ligament | Axial sesamoids | Distal metacarpus and proximal first phalanx |
| Palmar ligaments | Axial sesamoids | Corresponding abaxial sesamoids |
| Superficial digital flexor | Base of the medial epicondyle of humerus; cranial to humeral head of deep digital flexor muscle | Second phalanx by means of middle scutum |
| Deep digital flexor | Humeral head-medial epicondyle humerus Radial head-wide attachment to medial and caudal radius Ulnar-middle third of ulna | Base of third phalanx |
Sesamoidean Ligaments
The number and configuration of the sesamoidean ligaments are unique to the llama (Figures 58-3 and 58-4; see Table 58-1). The short sesamoidean ligaments attach the sesamoids to the corresponding first phalanx. The cruciate sesamoidean ligaments are short, compared with other species, and also attach the sesamoids to the corresponding first phalanx. The oblique sesamoidean ligaments attach the axial sesamoid bones to the palmar aspect of the opposite first phalanx. Interdigital intersesamoidean ligaments connect the two axial sesamoids to each other. Interdigital metacarpophalangosesamoidean ligaments link the axial sesamoids to both the distal metacarpal bones and the proximal first phalanx. Lastly, the palmar ligaments connect the axial sesamoids to the corresponding abaxial sesamoids. The sesamoidean ligaments are similar in the hind.3,4
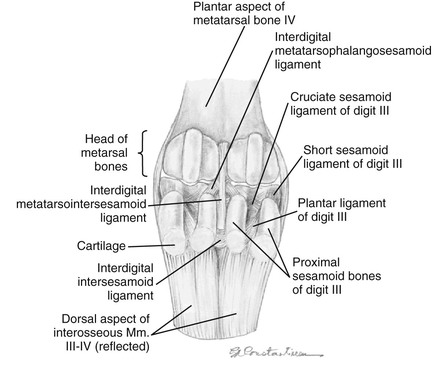
Figure 58-3 Left pelvic limb plantar aspect of the fetlock with the interosseous muscle (suspensory ligament) and sesamoid bones reflected to reveal the sesmoidean ligaments. (Courtesy of Dr. Gheorghe M. Constantinescu, University of Missouri. IN Constantinescu GM, Reed SK, Constantinescu IA: The Suspensory Apparatus and Digital Flexor Muscles of the Llama (Llama glama) 1. The Pelvic Limb, Int J Morphol 26(3):551-556, 2008)
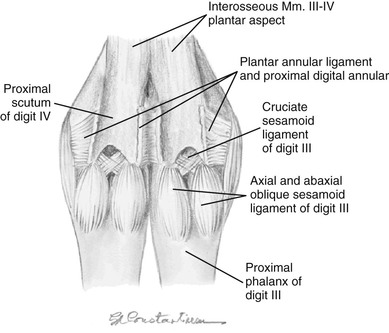
Figure 58-4 Left pelvic limb, plantar aspect of the fetlock joints with the superficial digital flexor and deep digital flexor tendons removed. (Courtesy of Dr. Gheorghe M. Constantinescu, University of Missouri. IN Constantinescu GM, Reed SK, Constantinescu IA: The Suspensory Apparatus and Digital Flexor Muscles of the Llama (Llama glama) 1. The Pelvic Limb, Int J Morphol 26(3):551-556, 2008)
Superficial Digital Flexor Tendon
In the forelimbs of the llama, the SDF muscle originates at the base of the medial epicondyle of the humerus, deep to the origin of the flexor carpi ulnaris muscle and cranial to the origin of the humeral head of the deep digital flexor muscle (see Table 58-1). As the SDF muscle courses distally, its fibers intermingle with the flexor carpi ulnaris muscle and the humeral head of the deep digital flexor muscle. The SDF muscle becomes tendinous (SDFT) just proximal to the carpus and runs through the carpal canal with the DDFT. Traveling through the carpal canal, each tendon has its own sheath. Distal to the carpus, the SDFT is continuous with the palmar fascia. The attached, thick palmar fascia fuses to the suspensory ligament on the lateral aspect of the fused metacarpals at the level of the origin of the suspensory ligament. The SDFT branches just proximal to the sesamoids into lateral and medial branches. The lateral and medial branches of the SDFT surround the corresponding DDFT at the level of the fetlock joint to form the manica flexoria (Figure 58-5).
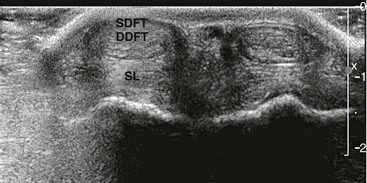
Figure 58-5 Transverse ultrasonographic image of the palmar aspect distal limb at the level of the fetlock. The branched superficial digital flexor tendon (SDFT) is surrounding the two branches of the deep digital flexor tendon (DDFT) at this level to form the manica flexoria.
The SDFT and the DDFT share a common distal flexor tendon sheath while crossing over the palmar aspect of the fetlock joint. The SDFT branches again distal to the fetlock to run palmarolateral and palmaromedial on each digit. At the level of second phalanx, the DDFT passes superficial to the SDFT, and the SDFT attaches to the second phalanx by means of the middle scutum. In the hindlimb, the superficial flexor muscle originates at the supracondylar fossa on the caudal aspect of the femur.3,4 The SDFT wraps around the calcanean tendon and has a branched attachment to the calcanean tuber. The SDFT anatomy and attachments in distal hindlimb are similar to those of the distal forelimb.3,4
Deep Digital Flexor Tendon
The origin of the deep digital flexor muscle has three heads in the llama: the humeral, radial, and ulnar heads (see Table 58-1). The humeral head originates on the medial epicondyle of the humerus and has two separate portions. The radial head originates with a wide attachment to the medial and caudal aspect of the radius. The ulnar head originates from the middle third of the ulna. Within the carpal canal, the heads fuse into one tendon. The DDFT splits into medial and lateral branches just proximal to the fetlock joint, distal to the location of the corresponding split of the SDFT. The DDFT is bound by the distal digital annular ligament as it passes over the second phalanx. At the level of the distal interphalangeal joint, the DDFT passes over the distal scutum and inserts broadly on the base of the third phalanx (Figure 58-6).3,4 The distal scutum is a thick cartilaginous plate connected to the second and third phalanges, which may serve a similar function as the absent distal sesamoid (navicular) bone.
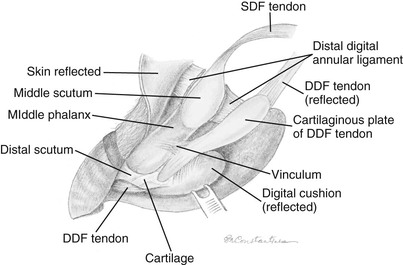
Figure 58-6 Left thoracic limb. Distal aspect of the foot demonstrating the insertions of the superficial digital flexor tendon (SDFT) and deep digital flexor tendon (DDFT). (Courtesy of Dr. Gheorghe M. Constantinescu, University of Missouri. IN Constantinescu GM, Reed SK, Constantinescu IA: The Suspensory Apparatus and Digital Flexor Muscles of the Llama (Llama glama) 1. The Thoracic Limb, Int J Morphol 26(3):543-550, 2008.)
In the hindlimb, the deep digital flexor muscle also has three heads: the lateral head, the medial head, and the underdeveloped caudal head (caudal tibial muscle). The caudal and lateral heads fuse proximal to the hock. The tendon of the medial head joins the other two distal to the sustentaculum tali. The distal aspect and attachments are identical to those of the forelimb.3,4
Lumbricalis Muscle
An important variation to be aware of in the llama, particularly when performing ultrasonography of the distal limb, is the presence of lumbricalis muscles (Figures 58-7 and 58-8). These are small muscles that lie on the palmar or plantar aspects of metacarpus or metatarsus between the digital flexor tendons. As the DDFT splits in the distal one third of the limb, the lumbricalis muscle becomes apparent. This muscle is spindle shaped and continues with a long and thin tendon that passes through the interdigital space on the axial surface of digits three and four, finally fusing with the lateral and medial branches of the lateral tendon of the common digital extensor in the forelimb and the long digital extensor in the hindlimb. In a published anatomic dissection of several llamas, variation in the number of lumbricalis muscles were seen between limbs and individual animals. Of the 8 thoracic limbs dissected, 6 limbs had one lumbricalis muscle, and 2 (belonging to the same animal) had two lumbricalis muscles per limb. In the 8 pelvic limbs dissected, 6 limbs had two lumbricalis muscles, and 2 limbs from the same animal had one muscle.3,4
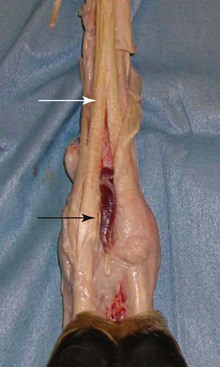
Figure 58-7 Photograph of the distal limb of the llama with the skin and superficial digital flexor tendon (SDFT) removed. The lumbricalis muscle (black arrow) is isolated from the deep digital flexor tendon (DDFT) at its bifurcation (white arrow).
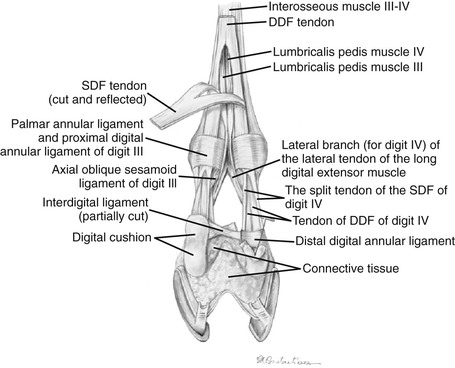
Figure 58-8 Distal left pelvic limb of the llama with the superficial flexor tendon reflected to reveal the emergence of the lumbricalis muscles at the splitting of the deep digital flexor tendon. (Courtesy of Dr. Gheorghe M. Constantinescu, University of Missouri. IN Constantinescu GM, Reed SK, Constantinescu IA: The Suspensory Apparatus and Digital Flexor Muscles of the Llama (Llama glama) 1. The Pelvic Limb, Int J Morphol 26(3):551-556, 2008.)
Anatomy of Diarthrodial Joints
Joint disease is found in camelids of all ages and may be caused by injury to the joint, bacterial infection, or degenerative osteoarthritis. To treat joint disease, it is imperative to know which anatomic communications or separations exist between joint compartments.2 Several important differences in joint communications are present in the llama compared with other species.2–12
Carpus
The camelid carpus has two rows of carpal bones as in other species (Figure 58-9). The proximal row contains the radiocarpal bone, intermediate carpal bone, and ulnar carpal bone, in addition to the accessory carpal bone caudally. The distal row consists of the second, third, and fourth carpal bones. The carpus is divided into three basic joint compartments: the radiocarpal, middle carpal, and carpometacarpal joints. Llamas have a greater percentage of communication between all three carpal joints than reported in cattle or horses.6,7 In llamas, the middle carpal and carpometacarpal joints always communicate, while the radiocarpal and middle carpal joints communicate about 32% of the time.2 Extensor tendon sheaths envelop the extensor carpi radialis, common digital extensor, and lateral digital extensor tendons along the dorsal surface of the carpus. The carpal canal contains the carpal sheath, a synovial sheath surrounding the SDF and DDFT. This sheath is separated from the carpal joints by the palmar carpal ligament. A high percentage (64%) of communication exists between the carpal sheath and the radiocarpal joint of llamas, which should be taken into account if sepsis of the radiocarpal joint exists.2
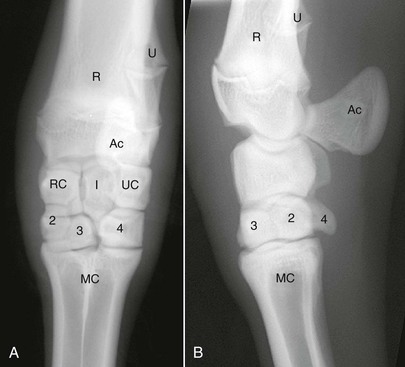
Figure 58-9 A, Dorsopalmar radiograph of the llama carpus. B, Lateromedial radiograph of the llama carpus. Ac, Accessory carpal bone; I, intermediate carpal bone; MC, metacarpus; R, radius; RC, radiocarpal bone; U, ulna; UC, ulnar carpal bone; 2, second carpal bone; 3, third carpal bone; 4, fourth carpal bone.
Joint Injection Landmarks.
The radiocarpal joint may be injected either lateral or medial to the extensor tendons, with the carpus flexed to open the joint space. The middle carpal joint may be injected lateral to the extensor tendons in a divot created between the intermediate carpal, ulnar carpal, and fourth carpal bones. The carpometacarpal joint may be injected on either side of the extensor tendons, by directing the needle axially toward a divot between C3, C4, and the third and fourth metacarpals. South American camelids (SACs) have a tough fibrous dorsal carpal pad that inhibits palpation of the extensor tendons. Therefore, inadvertent injection into the extensor tendon sheath may occur when performing arthrocentesis of the carpal joints.2
Tarsus
The camelid tarsus has three rows of tarsal bones (Figure 58-10). The proximal row consists of the talus and the calcaneus, the middle row contains the central tarsal bone, and the distal row has the first, fused second and third, and the fourth tarsal bones. The four principal joint compartments are the tibiotarsal, proximal intertarsal, distal intertarsal, and tarsometatarsal joints. In llamas, the tibiotarsal and proximal intertarsal joints consistently communicate, and 37% have communication between the proximal intertarsal and distal intertarsal joints.2 In addition, communication between the distal intertarsal and tarsometatarsal joints occurs about 68% of the time.2 All four tarsal joints communicate in 26% of the tarsi injected. Interestingly, when comparing paired tarsi within each llama, 23% have differences in tarsal joint communications.2 In llamas, the percentage of communication between the proximal and distal intertarsal joints and the distal intertarsal and tarsometatarsal joints is higher than previously reported in horses.2,8,9
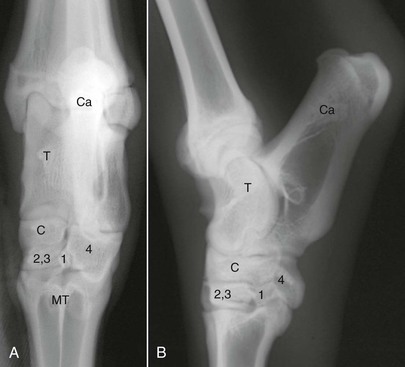
Figure 58-10 A, Dorsopalmar radiograph of the llama tarsus. B, Lateromedial radiograph of the llama tarsus. C, Central tarsal bone; Ca, calcaneus; MT, metatarsus; T, talus, 1, first tarsal bone; 2,3, fused second and third tarsal bones; 4, fourth tarsal bone.
Metacarpophalangeal or Metatarsophalangeal (Fetlock) Joint
The fetlock in camelids is formed by the third and fourth metacarpal or metatarsal bones, two pairs of proximal sesamoid bones along the palmar or plantar aspect, and two proximal phalanges (Figure 58-11). It has two compartments: medial and lateral. These compartments are separate in 94% of joints, and the remaining 6% may communicate along the proximal portion of the palmar or plantar joint capsule.2 Limitation of the communication to only the palmar or plantar aspect suggests that functional separation of joint compartments exists, even in those joints that communicate.2 Comparing the fetlock joints within each animal, differences were detected in lateral and medial compartment communications in 18% of metacarpophalangeal (MCP) and 4.5% of metatarsophalangeal (MTP) joints.2 The lack of communication between the medial and lateral compartments in llamas is similar to that in the dromedary camel and is the opposite of that in cattle.10,11
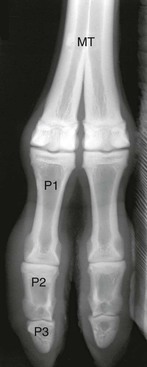
Figure 58-11 Dorsopalmar radiograph of a llama fetlock. MT, Metatarsus; P1, first phalanx, P2, second phalanx; P3, third phalanx.
Stifle
The stifle is one compartment in llamas that is similar to what has been described in sheep and dromedary camels.2,11–13 A single patellar ligament attaches the distal patella to the tibial tuberosity and the tibial crest.13 The proximal portion of the patella receives tendinous insertions from the quadriceps, including the vastus lateralis, vastus medialis, vastus intermedialis, and rectus femoris muscles, as well as fascia from the tensor fascia lata.13 Medial and lateral patellar retinacula help stabilize the patella within the trochlear groove.13 The llama also has lateral and medial menisci, cranial and caudal cruciate ligaments, and medial collateral ligaments as in other species. However, the llama does not appear to have a lateral collateral ligament; instead, the tendon of the fused peroneus tertius and extensor digitorum longus muscles provides lateral support to the joint.13
In other species, the stifle is composed of three different compartments that communicate variably. In horses, the lateral femorotibial joint is generally separate from the medial femorotibial and femoropatellar joint, whereas in cattle, communication between all three compartments occurs about 57% of the time.14,15 In llamas, all of the stifles that were successfully injected revealed communication between the three joint compartments.5 Infiltration of the freely movable tibial fat pad has been reported to inhibit penetration of the joint capsule.5
Orthopedic Diseases of Camelids
Alpacas and llamas are considered excellent orthopedic patients among large animals because of their quiet temperament, rapid bone healing, and relatively light body size. Besides fractures, many types of orthopedic diseases affect camelids, including angular and flexural limb deformities, osseous sequestration, joint injuries, and suspensory apparatus breakdown. Surgical correction has been described for many of these orthopedic diseases, with the outcome dependent on the animal’s age, body weight, duration of injury, and type of lesion.
Developmental Orthopedic Disease
Angular Limb Deformities
Angular limb deformities are fairly common in camelids. Mild bilateral carpal valgus is prevalent in both llamas and alpacas. A connection seems to exist between low vitamin D levels and the development of angular limb deformities in camelids, especially in the northwestern United States.16 Radiographic evidence of metaphyseal and epiphyseal flaring and physeal ectasia in animals having angular limb deformities is suggestive of hypophosphatemic rickets.17 Our clinical impression is that moderate to severe angular limb deformities secondary to hypophosphatemic rickets have become less common in recent years because of increased vitamin D supplementation during the winter months. Crias with dark hair coats are especially prone to developing low vitamin D levels because of reduced absorption of sunlight by the skin.18 Supplementation with vitamin D3 (1000 international units per kilogram [ IU/kg BW]) subcutaneously (SQ) every 60 days has been recommended for crias during the cloudy months of the year.19 Idiopathic adrenoleukodystrophy (ALD) does occur, and spontaneous correction is well recognized.
When spontaneous correction does not occur or angulation is severe, several surgical options for correction of angular limb deformities exist, depending on the degree of angulation, age of animal, and location. These options include hemicircumferential periosteal elevation, ulnar ostectomy, transphyseal bridging, and wedge or step ostectomies. Periosteal elevation of the concave side of the limb is recommended for mild to moderate angulation (<15 degrees) of the carpus similar to the technique used in horses.20 Anecdotal evidence suggests that this procedure should only be performed during the early phase of development (<3 months old). Camelids have complete ulnas (Figure 58-12) that fuse distally with the radial epiphysis. Partial ulnar ostectomy has been recommended for treatment of carpal valgus. A 2-centimeter (cm) segment of the ulnar bone is removed at a level immediately adjacent and proximal to the distal radial physis. For camelids less than 5 months of age with mild to moderate angulation of the carpus, periosteal elevation and ulnar ostectomy, done in combination, would be recommended. However, rapid periosteal bone formation may lead to premature fusion of the ulna and may complicate resolution of the angular deformity. For camelids over 5 months of age or those having moderate to severe angular limb deformities (>15 degrees), transphyseal bridging of the convex side of the limb appears to be a better treatment in conjunction with partial ulnar ostectomy (Figure 58-13). Unicortical screws are placed on either side of and parallel to the growth plate and then figure-of-eight orthopedic wires spanning around and between them. These wires develop tension across the physis and restrict the rate of growth on that side, thus allowing the opposite cortex to develop more quickly and straighten the angulation of the radius. Removal of the screws and wires should occur once the limbs have straightened. Failure to remove the screws and wires in a timely fashion may lead to overcorrection and significant carpal varus formation.
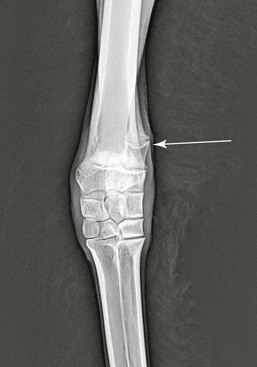
Figure 58-12 Complete ulna (arrow) in an 8-month-old male alpaca having bilateral carpal valgus deformities.
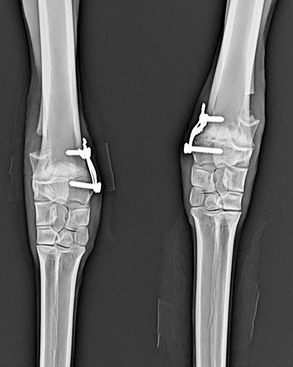
Figure 58-13 Transphyseal bridging and ulnar ostectomy has been performed in an 8-month-old male alpaca having bilateral carpal valgus deformities.
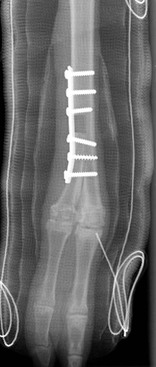
Deformities that are present after closure of the growth plate require step or wedge ostectomies to achieve correction (Figure 58-14). Surgical planning should include correction for rotation as well as angulation. Stabilization of the limb following ostectomy may be achieved via transfixation pin casts or plating.21,22 Arthrodesis may also be considered for angular limb deformities that have resulted in severe degenerative joint disease of an adjacent joint.23
Flexural Deformities
Congenital flexural deformities may be found in alpacas and llamas, most commonly affecting the fetlock, carpus, or tarsus. It is recommended that radiographs be taken of all affected joints to determine if concurrent bone malformation is present. Mild to moderate flexural deformities without underlying bone malformation should respond favorably to splinting or casting to correct the deformity. Severe flexural deformities may require transection of tendons or ligaments to achieve proper angulation of the joint.24,25 Results have been variable in these cases and have depended on which structures must be severed to achieve correction. Breakdown of the affected joint or hyperextension of the contralateral fetlock are potential complications following tenotomy or desmotomy for a flexural deformity. If underlying bone malformation exists or if tenotomy or desmotomy fails to correct the problem, corrective ostectomy or amputation of the affected limb may be considered. Amputation is feasible if only one limb is affected. Complications following limb amputation include contralateral limb breakdown and angular limb deformities secondary to increased weight bearing. Therefore, corrective ostectomy may be preferred in many cases.
Osseous Sequestration in Juveniles
Osseous sequestration is a recently recognized condition in juvenile camelids.26,26a Sequestration in horses and cattle has most commonly been associated with traumatic events leading to local cortical bone ischemia and bacterial invasion, secondary to loss of overlying periosteum and soft tissue viability.27,28 Interestingly, many bone sequestra diagnosed in camelids have not been associated with trauma or soft tissue wounds. Instead, these sequestra resemble ischemia to the bone, and bacterial emboli secondary to septicemia has been the most common hypothesis for the development of these lesions. Indeed, bacterial sepsis has been implicated as an etiologic factor of osteomyelitis in children and may be a factor in bone sequestration as well.29
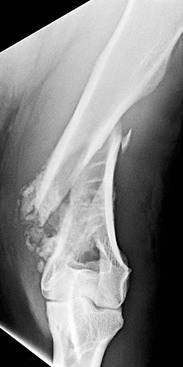
In a recent study, 36 camelids were affected by osseous sequestration.26a Of these 36 cases, 9 occurred in llamas and 27 were alpacas and the study included 22 intact males, 11 females, and 2 geldings. The median age of these animals was 7.5-months, 3.9-months, and 8.5-years, respectively. The appendicular skeleton, including the metacarpus, tibia, talus, femur, humerus, and proximal phalanx, was most often involved (Figure 58-15). Clinical signs included sudden onset of lameness and swelling in the affected limb (Figure 58-16). Open draining tracts are not typically seen, although purulent material may be present upon incising the skin in some animals.26,26a Surgical removal of the sequestrum often requires partial debridement of the bony callus to gain access to the sequestrum. In one animal, removal was unsuccessful because of the endosteal location of the sequestrum. In all other animals, sequestrectomy was considered successful and resulted in decreased lameness and swelling in the affected limb. Complications following sequestrectomy may include pathologic fracture through the sequestrectomy site if insufficient callus is present (Figure 58-17). Therefore, sequestrectomy may need to be delayed until maturation of the involucrum is evident radiographically or, in the case of large sequestra, until sufficient stabilizing callus has formed. Alternatively, additional means of stabilization or coaptation may be necessary to prevent pathologic fracture. The prognosis after squestrectomy is expected to be good with 83% of patients having recovered fully in one study.26a
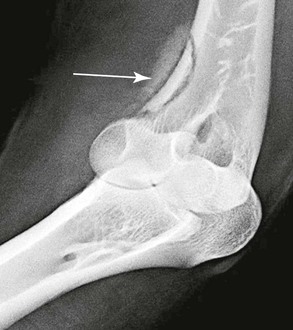
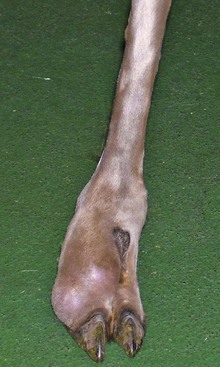
Figure 58-16 An alpaca exhibiting soft tissue swelling caused by subcutaneous infection and osseous sequestration of P1.
Joint Injuries
Septic Arthritis
Septic arthritis in camelids occurs mainly secondary to lacerations or traumatic injuries (Figure 58-18). Bacteremia in neonates may result in joint sepsis, although this cause does not appear to be as common in camelids as is found in foals. Because communication between all joint compartments may occur within the carpus and the tarsus of the llama and differences of joint communications may be found within each animal, it is important to determine the individual animal’s actual joint communications prior to treatment of septic arthritis.2 Therefore, each compartment of the carpus or the tarsus should be sampled for joint fluid cytology, injected with sterile contrast media for radiographic determination of joint communications in llamas, or a combination of both before instituting appropriate treatment.
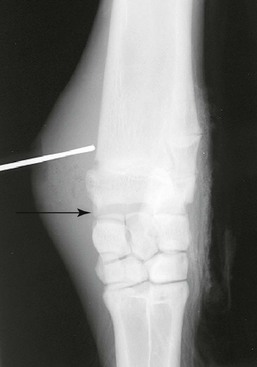
Figure 58-18 A metal probe has been placed in a wound over the medial aspect of the distal radius in a llama. Note the widened radiocarpal joint space (arrow). The radiocarpal joint did not communicate with the middle carpal joint in this llama.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


