3 Factors affecting feed intake
Knowledge of food intake is basic to the appropriate feeding of animals. Knowledge of daily nutrient requirements per se is of no value if the likely food intake of an animal is not known because the nutrients have to be “packaged” into a mass that the animal can consume within a 24 h period. Furthermore, underfeeding a horse in terms of feed mass, although not necessarily food nutrients, will mean that appetite will remain unsatisfied and, in the case of horses, this can lead to the development of undesirable stereotypic behaviors. Thus, it is essential to know the likely food intake of a horse or pony and also, to be aware of those factors that can moderate intake. Nowadays, obesity amongst the equid population is a problem of increasing significance and in order to limit or even control the intake of energy we must discover how to moderate this intake in a “welfare-friendly” fashion in order to maintain the health status of these animals. This chapter provides a summary of factors that are thought to influence intake (quality and quantity, palatability and appetite) as well as rate of intake. Hormonal regulation of feed intake is briefly discussed elsewhere (Chapter 2).
Quantitative intake
It is not known whether the controls of intake are the same for horses, ponies, different sexes, ages or breed types. However, we do know that breed size affects the rate of intake because of differences in the size of the harvesting (incisor arcade) and food processing features of the mouth (molar battery) that affect “handling time”. This time increases linearly with bite size and the maximum processing rate increased with body size (Fleurance et al 2009). These authors suggested that smaller breeds are more constrained when bite size increases. Furthermore, they discovered a significant effect (p < 0.0001) of the bite size × NDF interaction in ponies, which suggests that they are less well-adapted to dealing with high fiber feeds. It seems that when they encounter high fiber swards they limit their chewing activities rather than increasing their effort in handling.
Feedback mechanisms
Several feedback mechanisms appear to affect food intake by horses and these are detailed below.
Oropharyngeal monitoring
Ralston (1984) concluded that horses rely primarily on oro-pharyngeal and external stimuli to control the size and duration of an isolated meal. This conclusion was based on a study with ponies that were sham-fed but performed normal sequences of satiety behavior following a meal. However, only three ponies were used in the study and it is still possible that both metabolic and gastrointestinal inputs could moderate feeding behavior. The author considered that meal frequency is regulated by the presence and/or absorption of the products of digestion together with metabolic cues that may reflect body energy stores. However, the influence of the latter may be deemed weak in view of the fact that fat ponies tend to get fatter and remain fat. It is generally accepted (Cuddeford pers. observation) that equids are more sensitive to the feel, smell and taste of their food than are ruminants and thus, oropharyngeal monitoring must be more important in this species. This seems to be the case especially when offered clamp silage (Moore-Colyer & Longland 2000) or when fed straw. Dulphy et al (1997b) suggested that the organoleptic qualities of the latter may well limit its intake by horses.
Digestion end products
Ralston et al (1979) suggested a relationship between plasma glucose, subsequent meal size and rate of eating by ponies after a 3 h fast. After a 4 h fast, a control infusion of 2 of water had no effect on feeding behavior whereas an infusion of 2 liters of water containing 300 g of dissolved glucose decreased intakes of a pelleted diet 0 to 3 h post-treatment (Ralston & Baile 1982a). This probably represents a postabsorptive effect rather than a localized gastric response because of its duration. However, the tonicity of the glucose infusion solution could have affected fluid balance that, in itself could have had an effect. Ralston and Baile (1983) showed that reductions in intake occurred at times that would, in fact, reflect the postabsorptive state; 10 to 15 min for glucose and 4 to 6 h for cellulose. An intragastric infusion of 133 g of corn oil immediately after a 4 h fast, and before being given ad libitum access to a pelleted diet, did not affect consumption of the diet but did reduce subsequent intake by tripling the normal inter-meal interval. This could have resulted from metabolic feedback or a reduced rate of gastric emptying. Ralston and Baile (1982a) concluded that elevated levels of plasma glucose and insulin do not immediately generate satiety cues in ponies, although this view was based on glucose infusions via the jugular vein. Such an infusion would enable utilization of the metabolite and possibly, reduce effects at receptor sites. It is also probable that the glucose would have bypassed liver receptors as well. In this context it is worth noting that portal vein glucose sensors monitor glucose uptake from the gut and are involved in whole body glucose disposal, at least in rodents (Hevener et al 2001), dogs (Pagliassoti et al 1996) and pigs (Schmitt 1973). The signals generated affect the function of the liver and pancreas, tissues that are involved in glucose homeostasis (Burcelin et al 2001). Furthermore, signals enter the central nervous system to regulate functions such as feeding and satiety (Schmitt 1973). However, in the horse, it is hard to imagine that such an animal that has evolved essentially to ferment forage with the resultant evolution of volatile fatty acids (VFAs), will have a well-developed glucose sensing system in the portal vein.
Fermentation end products
Fermentation occurs throughout the horses’ gastrointestinal tract (GIT) releasing VFAs such as acetic, propionic, and butyric acid. Most are produced in the large intestine and, following absorption, are available in the liver where they can be used as a source of energy or in fat storage. It is conceivable that, like glucose, they may have a role in appetite regulation. Intragastric infusion of acetate (0.75 mmol/kg LW) caused ponies to increase feed intake (p < 0.05) by reducing the duration of the first inter-meal interval (Ralston et al 1983). Intravenous studies with short chain VFAs have not been undertaken and as a result, it is not known how circulating levels of these acids may affect voluntary dry matter intake (VDMI). Following a 4 h fast, above normal levels of acetate (1.0 and 1.25 mmol/kg LW) and propionate (0.75 mmol/kg LW) were introduced as a bolus into the cecum of ponies. VDMI was reduced in that the first inter-meal interval was prolonged (Ralston et al 1983). An intracecal infusion of 0.4 mol propionate per kg LW significantly (p < 0.05) increased VDMI by 1.075 relative to control values. In contrast, an infusion of 1 mmol propionate per kg LW significantly (p < 0.01) reduced by 0.22, the size of the first meal consumed without affecting subsequent feeding behaviors. Thus, there is some evidence to suggest local effects of VFAs within the GIT but the work was very limited. Furthermore, these animals were provided with complete pelleted diets that were fed ad libitum and consumed in 10/11 meals per day. However, the relevance of this type of research to the intake of an ad libitum forage-fed horse or pony is questionable.
Physical distention
The effect of gut fill on intake is equivocal. Intragastric infusions of kaolin had no significant effect on VDMI or feeding behavior of ponies when compared with control animals (Ralston & Baile 1982b). The authors suggested that stomach fill had no effect on intake although this ignores the fact that kaolin is inert and that it would not generate stimuli analogous to those produced by feed residues. Removal of 1.2 to 1.5 liters of cecal contents (equivalent to 0.22 of total cecal content of a 200 kg pony) also had no effect on subsequent feeding behavior of ponies (Ralston et al 1983). However, intra-gastric infusions of α-cellulose significantly (p < 0.05) reduced total VDMI after 3 to 18 h post-infusion (Ralston & Baile, 1982b). Both of these results need to be interpreted with care because removal of a small amount of digesta is unlikely to have had much effect on intake and, in the case of α-cellulose, it could have had a “fill” effect, a metabolite effect or both. In fact, Ralston and Baile (1982a) suggested that intragastric loads of nutrients generate satiety cues in ponies that are not related to the volume or bulk of the treatment.
Neutral detergent fiber (NDF) apparent digestibility and NDF content of feed are not reliable predictors of VDMI (r2 = 0.266) in horses (Cymbaluk 1990), suggesting that physical capacity of the large intestine is unlikely to limit feed intake. However, if fecal output were a determinant of VDMI then large intestinal capacity could have a role in controlling intake. The flow of digesta through the horse’s GIT is slowed but it is not limited by particle size in the same way as it is in ruminants since there is no analogous structure to the reticulo-omasal orifice. Thus, digesta cannot be retained in the horse’s GIT in the same way that it can in ruminants and therefore, there is less likelihood of “fill” negatively affecting intake.
The mean retention time (MRT) of feed residues in limit-fed ponies (17.5 g DM per kg LW/72 g DM per kg W0.75) given lucerne silage alone, enzyme-treated silage or the same silage substituted with 0.30 DM sugar beet pulp (SBP) was not significantly different (Murray et al 2009). The excretion curves for hay and oats were very similar when fed to 500 kg cold-blooded Norwegian trotters and there seemed to be no difference in MRT between the hay and oats (Rosenfeld et al 2006). Furthermore, barley, maize, and wheat were ground, pelleted, extruded, or micronized to create a total of 12 processed grains and then fed in combination with hay to the same horses in order to measure gastrointestinal retention times (Rosenfeld & Austbø 2009). Processing appeared to affect passage rates and compartmental retention times, but did not affect the overall MRT. However, the authors seemed unclear as to which was the time-dependent compartment in the model used. The foregoing suggests that very different diets/feeds may have similar MRTs and thus, their respective intakes would be unlikely to be affected by feed residue residency times in the GIT.
Feed factors
Feed type
Fresh forage
Archer (1973) was probably the first person to investigate the grass species preferences of horses and showed that the most palatable sward comprised a clover-rich mixture. Pasture varieties of perennial ryegrass were very palatable and as acceptable as timothy and cocksfoot. Tall fescue, crested dogs tail and wild white clover were other species found to be palatable for horses. Smith et al (2007) used the n-alkane technique to quantify the VDMI of horses grazing a clover-rich sward (dry herbage mass 1741 kg/ha, DM 230150 g/kg, crude protein [CP] 101150 g/kg DM, NDF 828150 g/kg DM and acid detergent fiber [ADF] 691150 g/kg DM), given no other feed. The estimated VDMI varied from animal to animal in the range 32 to 54150 g/kg/day or from 150 to 254 g DM per kg W0.75 measured over a 4-week period. Similar high intakes were estimated recently (Longland et al 2011a) in ponies (268 ± 52 kg) based on LW change over a 6-week period at pasture (CP 129150 g/kg DM, NDF 423150 g/kg DM). The ponies averaged a daily gain of 0.96 kg and VDMI ranged from 0.029 to 0.049 LW. Edouard et al (2009) measured intakes up to only 152150 g/kg W0.75 but claimed to be the first to show that sward height affects patch selection and ingestive behavior in horses independently of variations in the nutritive value of forages. Daily VDMI averaged 21 g DM per kg LW per day (100150 g/kg W0.75) and varied between 14 and 32 g DM per kg LW per day (66 to 152150 g/kg W0.75) for individual horses; these differences were not significant. The horses selected vegetative patches of grass that they could ingest fastest and thus their feeding behavior accords with the predictions of optimal foraging models as proposed by Stephens and Krebs (1986) for large ruminant herbivores. It seems that horses have the ability to ingest large quantities of fresh herbage DM well in excess of need and that assumed by the National Research Council (2007) in their recommendations provided there is a high herbage mass. Furthermore, sward height rather than herbage quality seems to strongly influence selection. Clearly, estimates of the VDMI of fresh forage vary widely (66–254 g DM per kg W0.75) and those horses that consume large quantities will become obese and thus prone to metabolic disease. The question remains as to why some horses are able to ingest so much more DM than others?
There is a paucity of information in relation to the VDMI of fresh forage by horses, but that which is available is detailed in Table 3-1. Dulphy et al (1997b) included data for 16 fresh forages (covering a range of NDF from 300 to 611150 g/kg DM) and obtained a mean intake figure similar to that for grass hays whose NDF content varied from 495 to 709150 g/kg DM. Published values for fresh forages obtained from the literature by Dulphy et al (1997b) varied between 90 and 117150 g/kg, with a mean of 105 g DM per kg W0.75, not that dissimilar from the data that they derived. As a result of these Institut National de la Recherche Agronomique (INRA) experiments, these authors suggested a probable fresh forage (grass) intake of 19–22 g DM per kg LW in the absence of a reliable means of predicting intake based on the classical parameters such as crude fiber (CF) and NDF as used for ruminants.
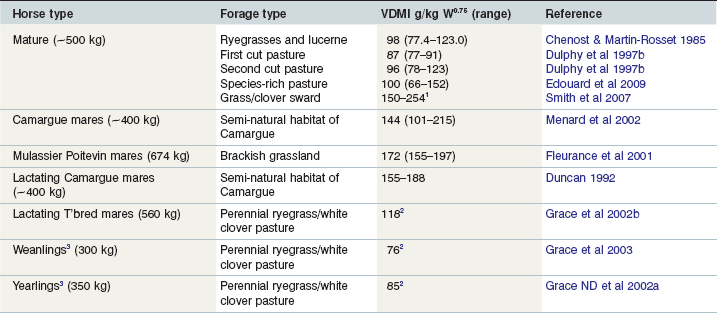
Forage intake work reported in New Zealand (Grace et al 2002a, b) with yearlings and lactating brood mares was based on fecal collection off the pasture and an in vivo assessment of pasture digestibility. The latter was done by cutting pasture and feeding it to confined horses in order to measure the apparent DM digestibility in vivo; VDMI by confined horses measured during a digestibility trial was lower than when grazing. The depression was 0.10–0.15 in stalled yearlings and 0.05–0.08 in corralled mares; both groups were offered about 0.30 excess herbage. This outcome illustrated the importance of grazing preference and its effect on VDMI (see earlier). The pasture intake by Australian stockhorse weanlings was measured differently by McMeniman (2000) in Australia using alkane markers but the results obtained (20–30 g DM per kg LW) were similar to those reported in New Zealand (~20 g DM per kg LW). Although the grass intakes by weanlings and yearlings in New Zealand met nutrient requirements, the intakes on the basis of W0.75 were rather low.
Conserved forage
Hay
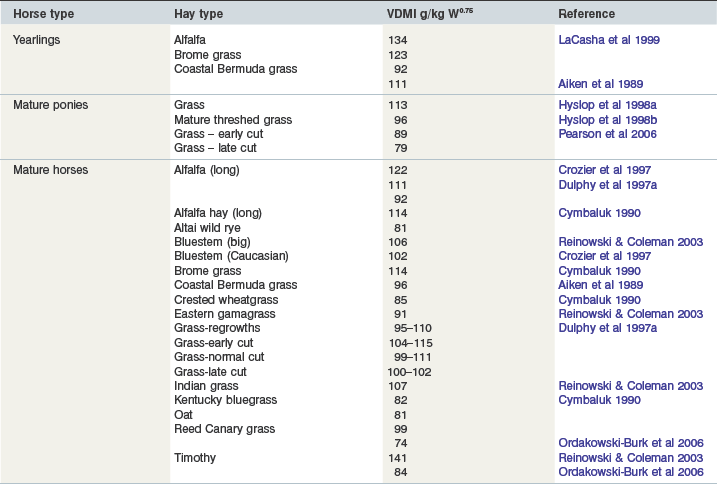
Cymbaluk (1990) compared a number of Canadian grass hays, fed ad libitum to horses, together with long alfalfa hay and dehydrated alfalfa pellets and it appeared that horses voluntarily consumed more alfalfa hay than grass hay (see Table 3-2). It seems to be a consistent finding that the intakes of legume hays are higher than that of grass hays and this may be for the reasons proposed by Minson (1990) to account for the higher intake of legumes by ruminants. He proposed that legumes had lower values for resistance to physical breakdown, percentage of cell wall, length-to-width ratios of fibers and energy required for mastication compared to grasses. In contrast, comparisons of the VDMI of chopped hays (legume and grass) by Arabian geldings (Crozier et al 1997) led to the conclusion that the higher intakes (p < 0.05) of legume hay, compared to grass hays were a reflection of its significantly higher (p < 0.01) dry matter digestibility (DMD). Overall, horses consume between 1.2 and 1.6 times more alfalfa hay than grass hay but the variation in intake between grass hays is not explainable although LaCasha (1999) has suggested that differences in cell wall composition may account for this variation between grass hays.
Ensiled forages
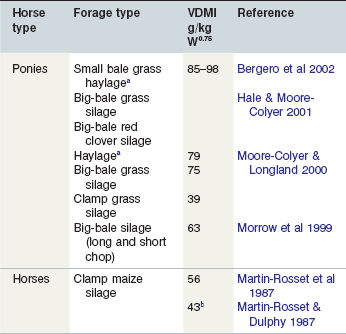
The VDMI of ponies fed limited amounts (1.65 kg DM per 100 kg LW) of grass-derived products (haylage, big-bale silage and clamp silage) were significantly (p < 0.05) different (Moore-Colyer & Longland 2000). The VDMI of clamp silage was less than 0.50 of that when the ponies were fed haylage, and hay VDMI was 0.80 of haylage VDMI. In view of the fact that the animals were limit-fed it would seem that intakes of the hay and clamp silage were probably limited by oropharyngeal factors such as palatability (for example silage fermentation characteristics including pH, VFA levels, DM, etc.) or, masticatory effort. Relative DM content of the hay, haylage, big-bale and clamp silage were 922, 676, 500 and 337150 g/kg; the haylage was preferred against the high DM hay but the VDMI of the clamp silage was low. DM content seems irrelevant in relation to intake and physical distention/gut fill could not have limited intake of the clamp silage as the ponies consumed 2.93 kg/day more of the big-bale silage. Similar, but lower, intakes of big bale silage containing either long or short chopped grass have been reported (Morrow et al 1999). It appears that the type of forage ensiled has a major impact on its subsequent intake by horses. For example, Hale and Moore-Colyer (2001) measured the VDMIs of big-bales of grass or red clover silages and compared them with that of grass hay. Ponies consumed significantly (p < 0.05) more red clover silage than hay, a finding in agreement with data presented in Table 3-2 that shows horses will eat more legume than grass-based forage; grass silage intakes were intermediate. The authors argued that maintenance of a constant bodyweight implied some degree of intake regulation although the short duration of the trial makes this conclusion uncertain. Low intakes of clamp grass silage (<0.50 of hay intake) by horses have been previously reported by others (McLean et al 1995). These low intakes were associated with significantly (p < 0.01) longer MRT compared to a hay diet; 51.7 h compared with 36.8 h using either chromium-mordanted hay or C32 alkane markers (McLean 2001). Since physical regulation of intake appears to be unimportant in the horse, DM content of the forage per se cannot account for these measured differences although intake of maize silage has been shown to fall as dry matter content falls (Agabriel et al 1982). However, the nature of the conservation process is usually less good in low dry matter silages and the fermentation characteristics of clamp silage have long been known to affect the VDMI of ruminants (for example see McCullough 1966) and perhaps they also affect the VDMI of horses. Ad libitum feeding of maize silage to horses resulted in very low intakes (43 g DM per kg1W0.75) and considerable refusals although conservation quality was considered satisfactory (Martin-Rosset & Dulphy 1987). This contrasted with ad libitum hay intakes in the same study of 99 g DM per kgW0.75. Recently, Ragnarsson and Lindberg (2010) limit-fed Icelandic horses (363 kg) a big-bale haylage (0.9 timothy grass, 0.1 meadow grass, DM 679150 g/kg) at the rate of 79 g DM per kgW0.75 with 0.05 refusals in the first collection period and none thereafter; this accords with the VDMI haylage values shown in Table 3-3. It follows from the above that horses will have a high VDMI when fed high DM haylages.
Dehydrated products
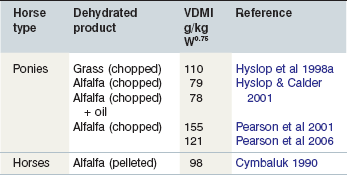
There is a large variation in the published intake values for high temperature artificially dehydrated forages (see Table 3-4). The alfalfa-based product used by the UK researchers had the same origin and yet, Pearson et al (2001, 2006) recorded much higher intakes and they suggested that this might be as a result of longer photoperiod during their trials. The parent material used for the UK product was young alfalfa and yet intakes of alfalfa hays (Table 3-1) were similar suggesting that stage of maturity of alfalfa has little impact on its intake by horses.
Cereal byproducts
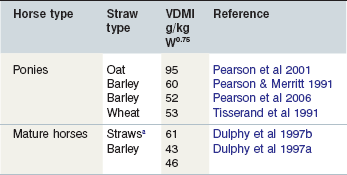
Dulphy et al (1997a) measured the VDMI of barley straw by horses to be approximately half that of grass hays (see Table 3-5). Pearson and Merritt (1991) had previously reported a similar situation in resting donkeys; 81 and 37 g DM per kg W0.75 per day respectively for hay and barley straw. However, the difference was not so great in resting ponies, 99 and 60 and surprisingly, the ponies appeared to be better able to compensate the poor quality of the forage offered. Pearson et al (2001) reported that the VDMI of oat straw by ponies was reduced by about 0.40 compared to when the ponies were fed ad libitum artificially dehydrated alfalfa. Similarly, Dulphy et al (1997b) reported a pooled intake value for undefined straws by adult light horses at maintenance that was rather lower than that reported for other dry forages. Thus, horses do not appear to be able to effectively compensate lower feed energy densities by eating more, although there is no evidence to support the view that intake is limited by the capacity of the GIT. Longer MRTs may inhibit intake although they may in themselves be a result of low intakes, furthermore, rate of elimination from the hindgut may have an impact on intake. Low intake of “cereal straw” may be influenced by its low palatability, a complex of organoleptic qualities including taste, structure, texture and smell. Surprisingly, results reported by Hyslop and Calder (2001) showed that the inclusion of approximately 0.50 oat straw in a short-chopped, dehydrated and molassed alfalfa diet had very little effect on total DMI, reducing it from 79 to 73 g DM per kg W0.75. This confirmed the findings of Hansen et al (1992) who showed that an inclusion of 0.50 wheat straw in a chopped alfalfa hay diet had little effect on VDMI (79 vs. 76 g DM per kg W0.75) in limit-fed (0.016 LW) horses although DMD was significantly (p < 0.01) reduced from 0.49 to 0.36. It seems that cereal straws have a VDMI half that of grass hays but they do not seem to depress intake of legume hay when included at 0.50.
Dulphy et al (1997b) observed that VDMI was not influenced by forage CP content although the very low CP content of cereal straws could inhibit microbial fermentation in the hindgut. Hyslop et al (1997) showed that the hindgut microflora of the pony were similar to those in the rumen in terms of degrading feeds and Van Soest (1994) noted that diets containing <80 g CP per kg DM depress the VDMI by ruminants through the slow rate of fermentation in the rumen and thus, slow reduction in particle size. However, as noted above, digesta outflow from the hindgut of the horse is not limited in the same way as that from the rumen. An alternative explanation for low intakes of straw could be that the rate of comminution (food processing) falls as the quality of the diet declines (Forbes 1988) so a poorer quality diet, such as straw, takes longer to eat. In addition, longer inter-meal intervals are required and if time spent feeding is an important feedback mechanism in intake control, then it is understandable that horses have reduced intakes of straw. However, it has been shown recently (Fleurance et al 2009) that handling time (mouth processing) increased linearly with bite size but it was not affected by the fibrousness of swards. Furthermore these authors showed that smaller animals (ponies) would be more affected by sward fibrousness than larger horses. Their response was not to increase handling time in a proportionate manner and thus it was hypothesized that compensation would be achieved by increased digesta passage rate thus allowing for greater intakes. This in part is supported by some comparative work with donkeys and ponies (Pearson et al 2001) that were fed ad libitum oat straw. The ponies appeared to consume more DM with shorter retention times in the GIT and with lower apparent nutrient digestibilities compared to when fed alfalfa. The exceptionally high oat straw intake compared to other straws may also have been due to a seasonal effect.
Concentrated feed (compound/cereal)
Horses are rarely allowed ad libitum access to concentrated feed. One study was reported (Cuddeford & Hyslop 1996) in which ponies were offered a high-fiber feed (NDF 418, CP 166150 g/kg) and consumed between 76 and 137 (mean 104) g DM per kg W0.75 which showed a great deal of variation in intake between ponies. A recent study (Henneke & Callaham 2009) investigated whether or not horses would self-regulate when given ad libitum access to a moderate energy-dense concentrate (C) and hay (H) in two trials. In a stall trial, total feed consumption began at 0.025 LW (0.01 C, 0.015 H), peaked at 0.0325 (0.025 C, 0.0075 H) and declined to 0.024 (0.021 C, 0.003 H) after a 21 day period. In a paddock-based trial, concentrate consumption peaked at 0.033 LW (22.45 kg/day) but eventually declined to 0.015 LW. Not surprisingly weight and condition score increased in both trials. Throughout these trials there was no evidence of colic or laminitis and the authors concluded that horses do have the ability to self-regulate the intake of both hay and concentrate after a suitable period of ad libitum feeding (~21 days) but some prefeeding, over a 2–5-week period, was necessary to allow the animals to adjust. There are insufficient data to allow firm decisions to be made about whether or not horses will self-regulate in the long-term and meet or exceed their needs. In the case of ponies, there is considerable risk in allowing them to “over eat” because of the concurrent risks of obesity and laminitis. However, it is clear that photoperiod (see below) affects the voluntary intake of ad libitum fed growing pony colts offered a complete pelleted diet (Fuller et al 2001). Long daylength resulted in increased intakes whilst short daylengths had the converse effect although responses were not immediate, taking 5 to 8 weeks to become evident.
Feed characteristics
Fiber and protein content
A comparative study conducted in France (Dulphy et al 1997a) with mature horses showed very similar VDMIs for different hays. Alfalfa, regrowth grass, early cut, normal cut and late cut grass hays were fed in two trials. The authors concluded that because the horses consumed the different hays in practically the same amounts, it would be difficult to define precise criteria whereby it would be possible to predict likely intakes of dry forages by horses. Furthermore, the horses did not increase their intake as the energy concentration of the feed declined. Ponies offered ad libitum mature, threshed grass hay (Hyslop et al 1998c) were able to consume relatively large amounts of dry matter, even though this hay was of very poor quality (DMD, 0.30). These results supported the view of Dulphy et al (1997a) that there is a poor relationship between VDMI and the cell wall content of hays. A further experiment (Hyslop et al 1998a) showed that although the hay used was of poorer quality than the other forages fed to mature ponies, more was consumed. The difference in intake was non-significant but the difference in NDF digestibility was (p < 0.01). Edouard et al (2008) have shown that forage digestibility declined as NDF increased and CP declined. NDF was a better predictor of DMD than CP for hays with a regression of DMD (%) = 100.3 − 0.78 × NDF (%) (r2 = 0.71, n = 38). These authors showed that the average VDMI of horses was unaffected by DMD or CP content but that VDMI appeared to decline slightly as NDF increased (DMI [150 g/kg LW] = 24.91 − 0.12 × NDF [%] [r2 = 0.10, n = 45]. An original discovery was that, by using mixed models that allowed for individual differences, as DMD declined, intake significantly increased. Similarly, as forage CP declined, VDMI increased. The authors concluded that most horses respond to a reduction in forage quality by increasing their VDMI as originally proposed by Janis (1976) but only until they reach a certain threshold as evidenced by the low VDMI of barley straw (0.5 that of grass hays, Dulphy et al 1997a).
Martin-Rosset and Dulphy (1987) and others (Pearson et al 2006) have suggested that variation in VDMI has little impact on the digestibility of the diet whereas other authors (Ott 1981, Cuddeford et al 1995) have shown a level of feeding effect. A study with Icelandic horses (Ragnarsson & Lindberg 2010) showed significant (p < 0.001) differences between digestibility coefficients for DM, organic matter (OM), NDF and ADF measured at two levels of intake, 47 and 79 g DM per kgW0.75. At first sight this contradicts the view of Martin-Rosset and Dulphy (1987) but examination of the French data showed that the CF digestibility of hay fed at 64 g DM per kg W0.75 was 0.551, at 86 g DM per kg W0.75 it was 0.487 and at 99 g DM per kg W0.75 it was 0.498; clearly there is a level of feeding effect. Although these data do not decrement equally, CF digestibility was consistently lower above maintenance (M) feeding levels; at 1.4M it was 6.4 units lower and 5.3 units lower when fed ad libitum. Furthermore, the Icelandic work revealed substantial individual variation in terms of the horses’ ability to digest fiber between feeding levels; NDF 0.012 to 0.102 (x = 0.065) and ADF 0.004 to 0.109 (x = 0.065). It should be no surprise that a limited intake of a quality feed will be associated with higher nutrient digestibility and a longer MRT; the refractory nature of straws, however, means that this scenario does not pertain to them.
A retrospective analysis of data obtained by the INRA and of values taken from published papers provided a data set for assessing VDMI of horses fed different dry forages (Dulphy et al 1997b). The pooled results for horses gave VDMIs for legumes (n = 12), grasses (n = 38) and regrowth grass hays (n = 7) of 108, 93 and 98150 g/kg W0.75 respectively; the only significant (p < 0.05) difference was between legume and grass hay. The authors concluded that voluntary intake of hay by horses was not influenced by either the CP, CF or NDF content of the forage. Multiple regression analysis yielded r2 values between 0.11 and 0.13; in contrast, r2 values for sheep calculated under similar conditions produced significant correlations of 0.78 to 0.84. Thus, it is impossible to reliably predict the intake of dry hays by horses based on forage analysis in contrast to ruminants, where effective equations have been derived and are based primarily on fiber content. Thus, Dulphy et al (1997b) have proposed that the voluntary intake of hays by housed, mature light horses will be in the range 19–22, 18–21 and 22–25150 g/kg LW for grass, regrowth grass and alfalfa hays respectively. These values may be regarded as conservative estimates of VDMI by horses.
In contrast to the above, a review (Lawrence et al 2001) of published studies that reported both NDF and VDMI in mature horses fed long stem grass hays ad libitum concluded that there was a good relationship between the two parameters represented by:
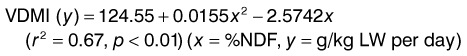 (1)
(1)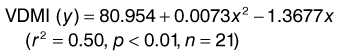 (2)
(2)More recently, Reinowski and Coleman (2003) used equation (2) to predict likely ad libitum intakes of different grass hays (see Table 3-2) and then measured the actual intakes of the hays; the intakes met or exceeded those that were predicted. There was a strong correlation between actual DMI and the NDF content (%) of the hays fed:
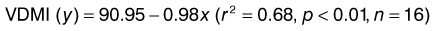 (3)
(3)Dulphy et al (1997a) were able to show a negative relationship between VDMI and plant NDF content as follows:
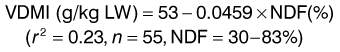
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


