Chapter 11 ANATOMY AND PHYSIOLOGY OF THE MARE
Comprehension of the anatomy and physiology associated with reproduction in the mare is essential for optimizing breeding management practices. This chapter provides an overview of structural and functional relationships associated with the reproductive system in mares compiled from previous and more recent reference texts and publications1–8 that is expected to serve as the foundation when considering the basis for various reproductive management regimens discussed in subsequent chapters.
TOPOGRAPHIC ANATOMY
The anatomy and relationships among various reproductive organs and supporting structures presented herein were compiled from several texts on veterinary anatomy in animals and the horse9,10 and reproductive biology in mares.1–48 Beginning with the central nervous system, the diencephalon of the forebrain is made up of three regions or glands (hypothalamus, hypophysis or pituitary gland, and epiphysis or pineal gland) that produce neural and endocrine signals to regulate development and maintenance of reproductive events. The hypothalamus is medially located and is the most ventral region of the diencephalon forming the lower parts of the lateral walls of the third ventricle; gonadotropin-releasing hormone (GnRH) is produced in this region of the brain. The pituitary gland is suspended below the hypothalamus by the infundibulum and lies in a recess on the floor of the cranium. The pituitary gland is composed of anterior, intermediate, and posterior lobes. The anterior lobe is associated with production of the gonadotropins, follicle-stimulating hormone (FSH), and luteinizing hormone (LH), which stimulate morphological (e.g., follicle and luteal development) and physiological (e.g., steroidogenesis) events within the ovaries, and prolactin, which is thought to primarily affect the mammary glands but may also be involved in seasonal reproductive changes. Thyroid-stimulating hormone (TSH) and adrenal corticotropic hormone (ACTH) are also produced by the anterior portion of the pituitary, in which the latter may also be produced to some extent by the intermediate lobe. Both hormones can have direct and indirect effects on reproduction, but they are primarily involved in growth, development of the central nervous system, and homeostasis. The posterior pituitary produces oxytocin, which is involved in morphological (e.g., smooth-muscle contractions) and physiological (e.g., endometrial prostaglandin production) events that affect the reproductive tract and mammary glands. Structural and functional abnormalities of the pituitary gland in horses (e.g., neoplasm or tumor) not only affect the reproductive organs directly but can commonly affect the adrenal glands (e.g., pituitary pars intermedia dysfunction, previously called Cushing’s disease) and thyroid and, therefore, affect reproduction indirectly.5,7. Finally, the pineal gland is a small, median body projecting dorsally from the brain stem behind an invagination of the roof of the third ventricle and is responsible for the production of melatonin that is also thought to play a role in seasonality.
The arrangement of the mare’s reproductive tract has been depicted and illustrated in several reference texts.1,2,8,10 The arrangement presented in Fig. 11-1 has been adopted from one of those texts1 to facilitate the following discussion. Overall, the reproductive tract is composed of the ovaries and tubular genitalia, associated vascular, lymphatic and nervous tissue, and suspensory ligaments. Most of the reproductive tract lies within the abdominal cavity and the remainder lies within the pelvic cavity. The bony architecture around the pelvic cavity is enclosed by symmetric halves of the hip bones (ilium, ischium, pubis), sacrum, and the last few tail vertebrae. Internally, the entrance to the pelvic cavity is represented by an inlet and outlet that can be demarcated by the position of the intravaginal transverse fold separating the vagina proper (cranial) from the vestibule (caudal). The cavity outlet may be smaller than the inlet, but the former expands during parturition as the wide sacrosciatic ligaments that border the outlet begin to relax in response to the placental hormone relaxin, especially as parturition approaches. Despite the distensible effect of relaxin on the pelvis, either the cavity inlet or outlet can represent major impediments during parturition as reviewed2,5,7,8 and discussed in Chapter 22.
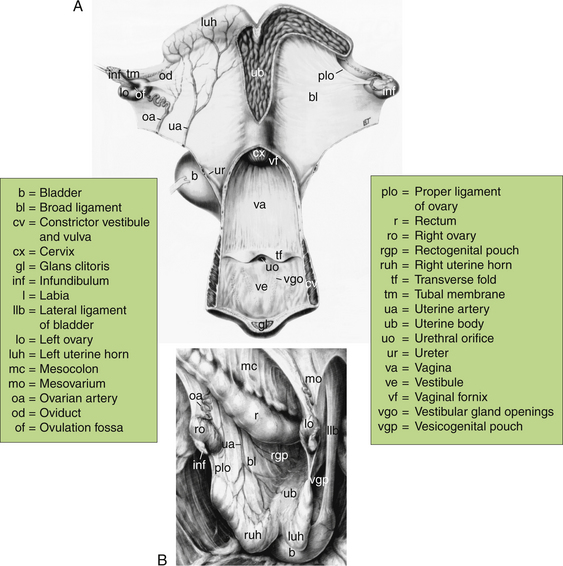
Figure 11-1 Dorsal (A) and lateral (B) views of the reproductive tract in the mare.
(From Ginther OJ: Reproductive Biology of the Mare: Basic and Applied Aspects, 2nd ed. Cross Plains, WI: Equiservices Publishing, 1992.)
Developmentally, the cranial portion of the reproductive tract is of mesodermal origin and the caudal portion is of ectodermal origin. The intravaginal transverse fold marks the convergence of these tissues from different origins (see Fig.11-1, A). Thus, the cranial portion of the reproductive tract includes the ovaries, oviducts, uterus, cervix, and vagina proper, and the caudal portion includes the vulva, which is often subdivided to include the vestibule, labia, and clitoris.
In situ size, shape, and placement changes of the reproductive tract are influenced by breed or type of horse (riding, draft, pony, miniature), season (ovulatory, anovulatory, transitional), age, reproductive status (puberty, nonpregnant, pregnant, post-partum, parity), body condition (weight), and health as well as whether the mare is in a standing position or sternal or lateral recumbency. More immediate changes are influenced by the extent of distention and activity of the colon, rectum, and urinary bladder. The ovaries and uterus are in intimate contact with the abdominal viscera and, therefore, may be suspended among the intestinal loops and appear Y-shaped or ride upon the viscera and appear T-shaped when viewed dorsally (see Fig. 11-1, A). Depending on the extent of suspension, the uterus may appear V-shaped when viewed from the side (see Fig. 11-1, B). The ventral aspect of the rectum and the dorsal aspect of the urinary bladder are in close contact with the uterine body. Consequently, the body of the uterus may be positioned laterally, away from midline, as a result of fullness of either the rectum or bladder. Thus, consideration of the physical relationships of the ovaries and uterus with the abdominal viscera and bladder is important when examining the reproductive tract using transrectal palpation or ultrasonic imaging techniques.11,12
STRUCTURAL ANATOMY
Suspensory Ligaments and Related Structures
The cranial portion of the reproductive tract, which is mostly in the abdominal cavity, is relatively fixed to the body wall by the broad ligaments.1–4,8,10 The broad ligaments are bilayers of serous membranes of the peritoneum that originate from the sublumbar region (approximately the third or fourth lumbar vertebra to the fourth sacral vertebra). The caudal portion of the reproductive tract in the pelvic cavity is retroperitoneal and lies within loose fascia and adipose tissue. Apart from their physical role, the broad ligaments have a functional role as corridors for blood and lymphatic vessels as well as nerves serving the reproductive organs. The broad ligaments are divided into areas with respect to the reproductive organs that they support. The mesometrium represents the majority of the broad ligament because it is supportive of the relatively large uterine horns and uterine body. The mesosalpinx is supportive of the oviducts, and the mesovarium is supportive of the ovaries. Portions of the mesosalpinx and mesovarium contribute to the ovarian bursa, an enclosed pouch into which the ovary can project. The lateral wall of the bursa is formed by the mesosalpinx, and the medial wall is formed by a fold of broad ligament with the proper ligament of the ovary on the free border of the fold. The proper ligament of each ovary attaches the caudal or uterine pole of the ovary to the cranial aspect of the respective uterine horn. Attachments of the various divisions of the broad ligaments are on the dorsal to lateral aspects of the tubular genitalia (see Fig. 11-1, B). Thus, for orientation purpose during transrectal palpation, the free surface of the uterine horns and body is ventral. To ensure proper assessment of the ovaries by transrectal palpation or ultrasonic imaging, it may be necessary to digitally manipulate the ovaries and position them medial to the broad ligament and free them from the mesovarium that extends over the medial and lateral surfaces.
Migration of portions of the digestive, urinary, and reproductive tract into the abdominal cavity and toward the pelvic cavity during embryonic and fetal development is associated with movement of respective peritoneal folds in which pouches form between suspending ligaments of respective organs. The rectum is dorsal and the bladder is ventral to the reproductive tract (see Fig. 11-1). Thus, the rectogenital pouch forms between the rectum and the genital organs, and the vesicogenital pouch forms between the bladder and the genital organs; both pouches project caudally into the pelvic cavity. These pouches are of clinical importance because they are entered surgically during colpotomy2,7,8 or during ultrasound-guided transvaginal manipulation of the ovaries7,8,11,12 and uterus13 as discussed in Chapter and 18. The bladder is suspended by two lateral ligaments and a ventral ligament that attaches to the pelvic floor. Transrectally, the lateral ligaments are readily palpable and are convenient for distinguishing the bladder from an advanced gravid uterus; the two lateral ligaments become extremely taut when the bladder is full.
Although the suspensory ligaments of the reproductive organs are the primary support for a gravid uterus, the ventral abdomen (i.e., linea alba and prepubic tendon) provides secondary support, especially during the later stages of pregnancy. Although the linea alba is mainly formed from the aponeuroses of the flank muscles, its caudal aspect combines with insertion tendons of the right and left rectus abdominis muscles to form a broad transverse plate, the prepubic tendon, which attaches to the pubis at the iliopubic eminence.9,10 With branching of the prepubic tendon, a relatively weak region exists cranial to the pubic brim between the two branches that connect to the pubis. Diagnosis, management, and prognosis associated with rupture of the prepubic tendon and other ventral abdominal ruptures or hernias, especially during advanced pregnancy, have been described.2,5
Blood Vessels, Lymphatics, and Nerves
The ovarian artery branches from the abdominal aorta and courses ventrolaterally along the cranial portion of the broad ligament (see Fig. 11-1). The artery eventually divides into uterine and ovarian branches; the uterine branch of the ovarian artery (cranial uterine artery) supplies blood to the oviduct and anterior portion of the uterine horn. The ovarian branch takes a more tortuous course within the mesovarium before dividing into several branches that spread out over the periphery of the ovary. The middle uterine artery branches and descends from the external iliac artery and is the main arterial blood supply to the uterus because of its large diameter, which expands even more during pregnancy. The uterine artery branches cranially and caudally as it approaches the dorsal mesometrial border of the caudal portion of the uterine horn and cranial portion of the uterine body. The vaginal artery (caudal uterine artery) originates from the internal pudendal artery and courses through the retroperitoneal tissue to supply blood to the caudal portion of the uterine body, cervix, and vagina. The vestibular branch of the internal pudendal artery supplies blood to the vagina proper, vestibule, and vulva. Essentially, veins draining the genital organs are satellites to the arteries, but the main venous drainage of the uterus is via the uterine branch of the ovarian vein. Unlike other species (e.g., bovids), the ovarian veins and arteries are not as closely opposed in the mare, which reflects the systemic route and uniqueness of how this species regulates the life span of the corpus luteum, especially during the mare’s first luteal response to pregnancy.1,14
In pregnant mares, there is a progressive increase in uterine weight and blood flow as gestation advances, stretching uterine arteries and making them susceptible to rupture or hemorrhage, especially during maternal strain and fetal movement and positioning associated with parturition, as discussed in more detail in Chapter 22. The right middle uterine artery is the most common site of vascular rupture, but the external iliacs and utero-ovarian (cranial uterine) arteries have also been known to rupture. Hemorrhage of a uterine artery may occur directly into the abdominal cavity or may be contained within the layers of the broad ligament; either condition is life threatening. Although blood emanating from the vulva may be a sign of uterine hemorrhage, it is more likely caused by rupture of varicose veins in the vagina since bleeding is not usually excessive and occurs during the latter part of gestation. Varicosities of the vaginal wall are most common in older mares, developing during estrus as well as pregnancy. Diagnosis, management, and prognosis associated with uterine hemorrhage and vaginal varicosities have been described.2,5,7,8
Lymph from the ovaries and cranial portion of the tubular genitalia passes to the lumbar aortic lymph nodes, whereas lymph from the caudal portion of the reproductive tract passes to the medial iliac lymph nodes and other lymph nodes in the pelvic cavity, all of which eventually reach the thoracic duct and empty into the venous system near the heart. The lymphatic system is a principal component of the uterine clearance mechanism for removing accumulated fluid in the uterine wall after estrus and particulate matter from the uterine lumen after insemination, embryo/fetal loss, and parturition. Blockage or damage of lymphatic vessels can alter optimal uterine clearance of fluid and debris; consequently, a hostile intra-uterine environment may ensue and jeopardize fertilization and pregnancy maintenance. Hence, oxytocin and/or low-dose administration of prostaglandin-F2α (PGF2α) are sometimes used in conjunction with or immediately after ovulation following insemination in mares with a history of subfertility that may be due to dysfunctional uterine clearance mechanism, as reviewed5,7,8 and discussed in Chapter 13.
Caudal Reproductive Tract (Perineum, Vulva, Clitoris, Vestibule)
The caudal portion of the reproductive tract is relatively stable within the pelvic cavity; therefore, gross shape and positional changes are minimal compared with the cranial reproductive tract that lies mostly within the abdominal cavity. The dimensions ascribed to the reproductive organs in this chapter relate primarily to excised organs taken from riding-type horse mares at necropsy and, therefore, may not be precisely representative of organ dimensions in situ.1,2,3,4,9
Perineum
The perineum or perineal region is broadly defined to include the external portions of the vulva and anus and surrounding area such that it may extend from the base of the tail to the ventral commissure of the vulva and, in some instances, to the dorsocaudal aspect of the udder. Any deviation from optimal conformation of the perineum (area surrounding the anus and vulva) may compromise the vulvar seal and result in the influx of air into the vagina (i.e., pneumovagina, or “wind-sucking”). Malconformation of the perineum may also result in the pooling of urine in the vagina (urovagina). Any of these conformational disfigurements of the perineum predispose the mare to acute or chronic inflammation of the vagina, cervix, and endometrium, thus affecting fertility. The degree of malconformation of the perineum and management (e.g., Caslick’s vulvoplasty operation) have been described.2,5,7,8
Vulva
The vulva, which usually includes the labia (vulvar lips) and clitoris, is the most caudal portion of the reproductive tract and is considered the first line of defense to protect against contamination of the uterus. The vulvar cleft is represented by a vertical slit (approximately 12–15 cm in length), with pointed dorsal and rounded ventral commissures, and marks the external orifice of the urogenital tract, which is controlled by the striated vulvar constrictor muscle that runs along either side of the length of the vulvar lips. The mucocutaneous junction of the labia demarcates the aglandular mucous membrane from the glandular pigmented skin. The gross appearance of the vulva/labia can be influenced by the stage of the estrous cycle as presented in Table 11-1. Approximately two thirds of the vulva extends caudoventrally over the ischial arch; therefore, safe passage of the forearm, pipette, or other instruments into the vestibule and vagina is at an upward angle.
Table 11-1 Tactile and Visual Changes of the Tubular Genitalia Using Various Instrumentation and Direct Sensation and Observation During the Estrous Cycle in the Mare
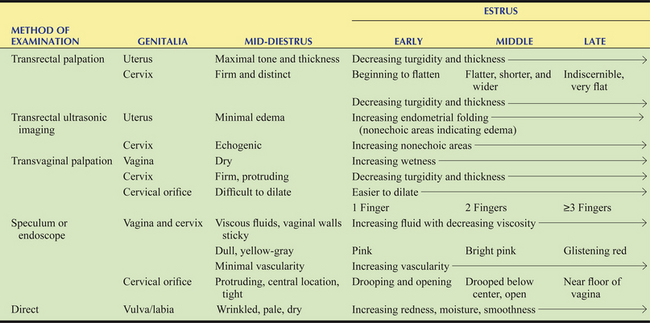
Adapted and summarized from Ginther OJ: Reproductive Biology of the Mare: Basic and Applied Aspects, 2nd ed. Cross Plains, WI: Equiservices Publishing, 1992.
Clitoris
The clitoral glans appears wrinkled and creased and is located in a protective cavity or pouch (clitoral prepuce) at the ventral commissure of the vulva (see Fig. 11-1, A). The clitoral retractor muscle and the vulvar constrictor muscle are responsible for the natural inversion of the labia and exposure of the clitoris (“clitoral wink”) during urination or during behavioral estrus. It can be exposed manually for examination and swabbing of its various sinuses (three to five) to test for harmful and contagious bacteria such as Taylorella equigenitalis responsible for equine metritis and Klebsiella pneumoniae and Pseudomonas aeruginosa as reviewed.2,5,7,8
Vestibule
The vestibule generally occupies a median position in the pelvic outlet and extends from the external vulvar lips to the transverse fold (approximately 10–12 cm in length) that often lies over the urethral orifice, caudal to the fold (see Fig. 11-1, A). In young, maiden mares, the fold may be partial or complete, forming the hymen, which may temporarily restrict entrance into the vagina until broken by the penis or manually by palpation. The mucous membrane of the vestibule has rows of papilla in the ventral and lateral walls that mark the opening of minor vestibular glands (see Fig. 11-1, A); striated muscles and the vestibular bulb (mass of erectile tissue) are associated with the lateral walls only.
Cranial Reproductive Tract (Vagina, Cervix, Uterus, Oviducts, Ovaries)
Vagina
The vagina proper (approximately 20–35 cm in length) in non-pregnant mares generally occupies a median position in the pelvic inlet while the remainder of the cranial portion of the reproductive tract lies in the abdominal cavity. The vagina usually exists as a collapsed lumen that is highly distensible in length and width to accommodate the penis or forearm during natural or artificial insemination and foal at parturition. The vagina proper is thin walled and aglandular but does receive secretions from the cervix, and, therefore, its gross appearance can be influenced by the stage of the estrous cycle (see Table 11-1). The vaginal fornix at the cranial aspect of the vagina represents an annular cavity in which the caudal portion of the cervix projects (see Fig. 11-1, A and Fig. 11-2). The mucosa is normally pale pink but darkens with prolonged exposure to air during vaginoscopy (see Fig. 11-2). During transvaginal ultrasonic imaging of the reproductive tract, it is the cranial wall of the vaginal fornix that is in contact with the face of the ultrasound transducer as well as the surface that is penetrated to enter the rectogenital pouch during an ultrasound-guided needle approach to the ovaries or uterus and that is incised during colpotomy.
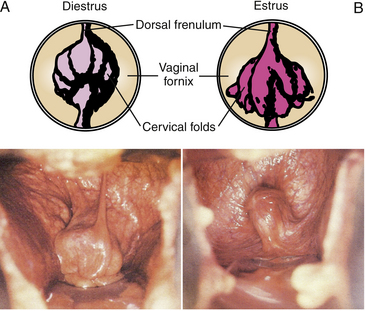
Cervix
The cervix is a sphincter-like muscle (5–8 cm in length and 2–5 cm in diameter) that protrudes caudally into the vaginal fornix (2–4 cm in length) and is the third and final line of defense to protect against contamination of the uterus (see Figs. 11-1 and 11-2). It has a thick wall of smooth muscle that is rich in elastic fibers and that changes in tone (turgid vs. flaccid), size, color, and secretions depending on whether the hormonal milieu is dominated by progesterone during diestrus and pregnancy (see Fig. 11-2, A) or estrogen during estrus (see Fig. 11-2, B) as indicated in Table 11-1. The cervix is arranged in longitudinal mucosal or cervical folds that are continuous with endometrial folds of the uterine body and horns (see Fig. 11-1, A). The intravaginal portion has a lobed appearance as a result of extension of the cervical folds into the vaginal fornix (see Figs. 11-1, A, and 11-2). A dorsal or ventral fold may continue onto the floor of the fornix as a distinct frenulum. A unique clinical feature of the longitudinal folds of the cervix is that they allow relatively easy access to the uterine lumen by digital or tactile dilation, especially during estrus, compared with domestic species with cervical rings (e.g., bovids). Unlike the aglandular vagina, mucus-producing cells of the cervix secrete copious amounts of mucus. The hormonally timed changes in cervical tone and its secretions are essential physical and physiological barriers to protect the uterus and facilitate uterine clearance, delivery, and transport of semen, fertilization, pregnancy development and maintenance, and parturition. Physical examination of the cervix can be done by using ultrasonography, palpation, a speculum, or endoscopy. Cervical dysfunction attributed to congenital defects or physical trauma and consequential effects on fertility have been reviewed.2,5,7,8
Uterus
The uterus consists of a uterine body and two uterine horns and is classified as a simplex bipartitus uterus (see Fig. 11-1). General dimensions in non-pregnant mares are that the length and diameter of the uterine body are relatively shorter (~18–20 cm in length) and larger (~8–12 cm in diameter) compared with longer (~20–25 cm in length) and smaller (~1–6 cm in diameter) uterine horns. The uterine body is continuous with the cervix and lies mostly in the pelvic inlet and abdominal cavity. Two uterine horns diverge from the uterine body and are joined by the intercornual ligament at the corpus-corneal junction; a short uterine septum marks the internal bifurcation (see Fig. 11-1, A). The corpus-corneal junction is important clinically because it represents a structural and functional region within each uterine horn where the early embryonic vesicle becomes “fixed” and readily detected by ultrasonic imaging to evaluate its status. Structurally, the degree of curvature or flexure of the uterine horns near the corpus-corneal junction (see Fig. 11-1, B), the increase in uterine tone (i.e., turgidity), and the expansion of the conceptus act in concert as physical impediments to trap the previous mobile vesicle in the caudal aspect of either the left or right uterine horn. The cessation of embryonic mobility that occurs 15–16 days after ovulation, defined as “fixation,” has been reviewed,1,2,11,12 and is discussed in Chapter 19.
Peripherally, the uterus is enclosed by visceral peritoneum—the perimetrium—and is continuous with the suspending portion of the broad ligament—the mesometrium. As discussed earlier, the mesometrium of the broad ligament attaches to the dorsal border of the uterine horns and the lateral aspects of the body and cervix (see Fig. 11-1, B).
The uterine wall consists of longitudinal (outer) and circular (inner) smooth muscle layers with a vascular layer in between, which makes up the myometrium (Fig. 11-3, A). The outer muscle layer and vasculature are continuous with the mesometrium. The myometrium is, in part, responsible for mobility of the early embryonic vesicle around the extent of the uterus and the force to expel the fetus at parturition.
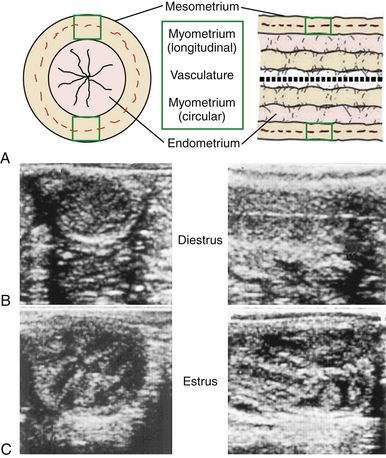
The intrauterine environment consists of a gland-free core of connective tissue arranged in longitudinal folds (12–15) and covered by a reddish-brown mucosa, the highly glandular endometrium. The lumen of the uterus is essentially collapsed, in which the endometrial folds are in close apposition but can be easily dilated by air or fluid. Ultrasonic morphology of the endometrium appears relatively homogeneous during diestrus (see Fig. 11-3, B) compared with heterogeneous during estrus (see Fig. 11-3, C). Changes in the ultrasonic echogenicity of the endometrium during the luteal and follicular phases are due to the degree of edema associated with the endometrial folds during progesterone and estrogen dominance (see Table 11-1), respectively, which has clinical importance in the timing of ovulation and insemination discussed in Chapter 12.
The endometrium is highly glandular with prominent openings on the surface toward the lumen; extreme branching and coiling of the tubular structures exist below the surface. Glandular changes are considered a prime indicator of uterine health when assessing uterine tissue biopsy specimens.2,5,7,8 Typically, histological assessment of glandular changes of an endometrial biopsy includes periglandular fibrosis, cystic dilation of glands, and glandular necrosis. In addition to biopsy score, microscopic evaluation of the endometrium may include characterization of lymphatic lacunae, endometrial atrophy, and cellular infiltration. Clinically, endometritis—acute or chronic inflammation involving the endometrium—is generally considered a major cause of reduced fertility in mares as reviewed2,5,7,8 and discussed in Chapter 13.
Oviducts
The tortuous oviducts (~20–30 cm in length) extending beyond the cranial tips of beach uterine horn are sometimes called uterine tubes or salpinges (see Fig. 11-1). There are three divisions extending from the ovary to the tip of the uterine horn: (1) funnel-shaped infundibulum with finger-like fimbriae, (2) expanded ampulla, and (3) narrow isthmus. Irregular fimbriae are present along the margin of the infundibulum, in which a portion of them are attached to the cranial pole of the ovary to form the cranial edge of the ovulation fossa. Remaining fimbriae and the infundibulum are free to spread over the ventral aspect of the ovary and cover the ovulation fossa.
The uterotubal junction marks the convergence of the oviduct and the uterine horn and, clinically, is sometimes the deposition site for sperm during low-dose intrauterine insemination as discussed in Chapters 14 and 15. Oviductal morphology is similar to the uterus (see Fig. 11-3, A
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


