Suborder Serpentes (i.e., the snakes) suborder Lacertilia (i.e., the lizards) suborder Amphisbaenia (the worm-lizards)
More recently the order Squamata has been divided into the:
Suborder Iguania (containing the Agamids, Iguanids, Chameleons and New world lizards)
Suborder Scleroglossa which itself is then split into the following infraorders
Anguimorpha (Monitor lizards, Heloderma lizards, slow worms etc.)
Amphisbaenia (worm lizards)
Gekkota (the geckos)
Scincomorpha (skinks, whiptail lizards and European Lacertid lizards)
Serpentes (the snakes)
Gavialidae (the gharial)
Alligatoridae (the alligators and caimans)
Crocodylidae (the crocodiles)
SNAKES
Like the bird, the snake has no diaphragm, so no separate thorax and abdomen. Instead it has a coelomic, or common, body cavity.
Musculoskeletal system
All true snakes have no limbs. This distinguishes them from species such as the slow worm, which is actually a lizard with vestigial limbs. There are some remnants of limbs in one or two of the older evolutionary species of snake such as the boid family (pythons and boa constrictors). These can possess vestigial pelvic remnants, having claw-like spurs either side of the vent representing the hind limbs.
The snake skull possesses a small cranial cavity containing the brain and a large nasal cavity. The maxilla has 4 rows of teeth, two on either side. The mandible has the more normal two rows of teeth (Figure 17.1). The teeth vary somewhat between the genera. The more commonly seen non-poisonous species such as the colubrid family (containing the kingsnakes and rat snakes) and the boid family, have simple, caudally curved, peg-like teeth. Some of the more poisonous species have specialist adaptations. Rattlesnakes, for example, have hinged, rostrally situated fangs which swing forward as they strike. All teeth are replaced as they are lost, including fang teeth in poisonous species. It is worth mentioning that owners (other than zoos) of poisonous species of snake, such as pit vipers and rattlesnakes, must be licensed and registered in the United Kingdom under the conditions of the Dangerous Wild Animals Act of 1976.
Figure 17.1 Intraoral view of a young Burmese python showing the four upper and two lower rows of teeth, as well as the glottal tube and tracheal entrance, rostral to which is the tongue sheath.
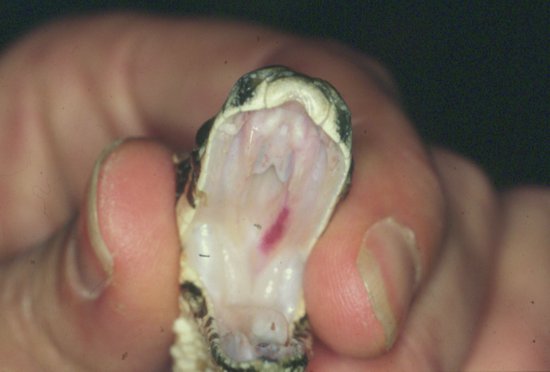
The anatomy of the snake’s head has a number of adaptations that allow it to swallow large prey. In all snakes, the two halves of lower jaw are loosely held together rostrally and the mandibular symphysis can separate. In addition, the snake has no temporomandibular joint. Instead it possesses a quadrate bone, which articulates between a mandible and the skull and allows the mandibles to be moved rostrally and laterally, ‘dislocating’. The maxilla also hinges only loosely with the rostral aspect of the cranium, so allowing the nose of the snake to be raised, increasing the oral aperture.
The skull articulates with the atlas vertebra via a simple joint containing only one occipital condyle, rather than the mammalian two. The coccygeal vertebrae (those caudal to the vent) are the only vertebrae with no ribs attached. Instead they have paired, ventral, haemal processes between which the coccygeal artery and vein run. This vein can be used for venipuncture both for sampling and for intravenous injections. The site for this is one third the distance from the vent to the tail tip on the ventral aspect.
The ventral scales, known as scutes or gastropeges, overlie the muscular casing of the snake’s torso. This muscle is segmental and supplied by intervertebral nerves. It is by alternately contracting and relaxing these segmental muscles that the snake can propel itself across the ground, the caudal edge of each ventral scute providing friction.
A very few species of snake, such as the glass snake, exhibit autotomy. That is, they will shed their tail if roughly handled or caught by a predator. They will regrow their tail later.
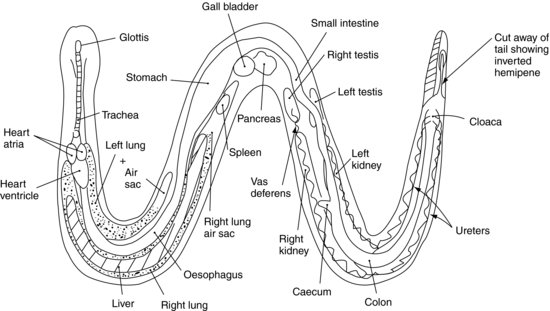
Respiratory system (Figure 17.2)
Upper respiratory system
The nostrils are paired and open into the roof of the mouth. Snakes, like all reptiles other than crocodilians, do not have a hard palate. When the mouth is closed, the internal nostrils are positioned directly above the entrance to the trachea. This is guarded by the glottis. An epiglottis may be present in vestigial form, but there is often fusion of the cartilages here to form a glottal tube. This tube is rigid enough to withstand the pressures placed upon it when the snake is swallowing whole prey. At rest the glottis is held closed, only opening when the snake breathes. The glottis then opens into the trachea which, in the snake family, is supported by C-shaped cartilages similar to those of the cat and dog.
Lower respiratory system
In the majority of colubrid species, such as rat snakes and kingsnakes and some Viperidae, the right lung is the major lung, the left having regressed to a vestigial structure. The vestigial left lung is often replaced by a vascularised air sac and so can take part in gaseous exchange. In the evolutionarily older species such as the Boidae there are two lungs.
The trachea bifurcates at the level of the heart. The lungs occupy the first half of the middle third of the body of the snake. As there is no diaphragm, inspiration is purely due to the outward movement of the ribs and intercostal muscles. This is aided by elastic tissue present within the lung structure, which allows the lungs to expand and recoil. Expiration is facilitated by contraction of abdominal and intercostal muscles, and the elastic recoil of the lungs themselves. A ‘tracheal lung’ is often present as an outpouching of the lining of the trachea from the open part of the C-shaped cartilages. This is thought to aid respiration when main lungs are being compressed during the swallowing of large prey items.
The stimulus for respiration is a lowered partial pressure of oxygen, rather than an increase in the partial pressure of carbon dioxide, as is the case in mammals.
Digestive system
Oral cavity
The tongue sits in a basal sheath at the rostral end of the oral cavity just in front of the glottis and can be pushed out through the lips even when the mouth is closed through the labial notch. The tongue is bifid (split into a forked end) and is used to catch odours on its moist surface. These are then pushed into the roof of the mouth into the vomeronasal organ. The vomeronasal organ is connected to the olfactory region of the brain and is a primitive but effective pheromone and scent detector. The oral cavity contains salivary glands which are stimulated to release saliva during mastication. The mouth is normally free of saliva at other times.
The oropharynx passes on into the oesophagus, which is an extremely distensible muscular tube travelling ventral to the lungs and entering the stomach in the second half of the middle third of the snake’s body.
Stomach, associated organs and intestine
The stomach is a tubular organ, populated with compound glands secreting both hydrochloric acid and pepsin (mammals having two separate cells) and separate mucus secreting glands. There is no well-defined cardiac sphincter. The majority of the digestive process occurs in the stomach and is continued by the small intestine. The only substance which cannot be digested is the hair of the prey, known as the ‘felt’, which is passed out in the stool.
The stomach empties into the duodenum, which is poorly defined from the jejunum and ileum. The spleen, pancreas and gall bladder are found at the point where the pylorus empties into the small intestine. Some snakes have a fused splenopancreas. The gall bladder is found at the most caudal point of the liver, which is an elongated structure extending from the mid-point of the lungs to the caudal stomach.
The small intestine empties into the large intestine, which is distinguished from it by its thinner wall and larger diameter. In the Boidae there may be a caecum at this junction.
The large intestine empties into the coprodeum portion of the cloaca, which, as with birds, is the common emptying chamber for the digestive, urinary and reproductive systems.
Urinary system
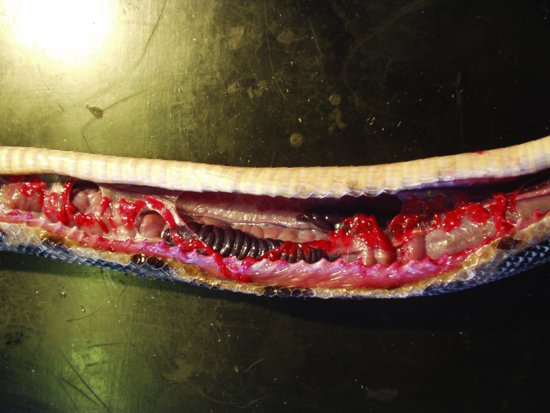
There are paired elongated kidneys (Figure 17.3), situated in the distal half of the caudal third of the snake’s body, attached to the dorsal body wall. The right kidney is cranial to the left and both has a single ureter each which travels across their ventral surface to empty into the urodeum of the cloaca, caudal to the proctodeum. There is no urinary bladder in snakes. The caudal portions of the kidneys in male snakes are the ‘sexual segments’, enlarging during the breeding season as they provide seminal fluid.
Figure 17.3 Postmortem showing the elongated lobular right and left kidneys of a rainbow boa constrictor (Epicrates cenchria).
Renal physiology
As with the majority of reptiles, snakes are uricotelic, that is, like birds their primary nitrogenous waste product is not urea but uric acid. This compound is relatively insoluble allowing conservation of water. This is particularly important for reptiles, as they have no loops of Henle in their kidneys; therefore, they cannot create hypertonic urine as mammals can. To further conserve water, urine in the urodeum portion of the cloaca can be refluxed back into the terminal portion of the gut where more water reabsorption can occur. If the reptile becomes dehydrated, or renal blockage or infection occurs, then excretion of uric acid is reduced and can lead to gout as is seen in birds.
Cardiovascular system
Heart
The heart is three-chambered, with two atria and a common ventricle situated within the pericardial sac, but it despite this functions as a four chambered organ. The heart lies in the caudal half of the proximal third of the snake’s body, and is mobile, to allow the passage of large food items through the oesophagus above it. There are two cranial venae cavae and one caudal vena cava entering via the sinus venosus (a narrow tube leading to the right atrium from which it is separated by the sino-atrial valve).
Blood vessels
The snake family has paired aortas. They exit one from each of the two sides of the single ventricle of the heart and then fuse into a single abdominal aorta. The pulmonary artery that leads to the lung(s) also arises here.
As with birds, snakes have a renal portal system. The blood supply from the caudal portion of the snake in the coccygeal artery splits into two and can enter the renal circulation or may bypass it via a series of valves. This is important when administering drugs which are nephrotoxic, or which may be excreted by the kidneys, as it means they might be concentrated there. They should therefore be administered in the cranial part of the snake.
There is also a hepatic portal system from the intestine to the liver. A ventral abdominal vein lies in the midline, just beneath the ventral abdominal musculature, and must be avoided when performing surgery.
Two external jugular veins run just medial to the ventral cervical ribs, and may be reached to place catheters for intravenous fluid administration via a surgical cut-down procedure. The ventral tail vein has already been mentioned and is useful for venipuncture for blood collection.
Lymphatic system
There are no specific separate lymph nodes as seen in mammals, a situation similar to birds. Instead, as with birds, there are discrete accumulations of lymph tissue within most of the major organs, particularly the liver and intestines. There is also a spleen, as mentioned above, which has loosely arranged red and white pulp. Lymphatic vessels are found throughout the body. A lymphatic sinus, for example, runs the length of the snake just ventrolateral to the epaxial musculature immediately below the skin surface on either side of the body. This may be used for small volumes of fluid administration. In the walls of many of the lymphatic vessels there are muscular swellings known as ‘lymph hearts’ which aid in the return of the straw-coloured lymphatic fluid back to the true heart.
Reproductive system
Male (Figure 17.2)
The paired testes lie intracoloemically (within the coelom or common body cavity, as snakes, like most reptiles, have no diaphragm and so no separate thorax and abdomen; rather they have only a common coelomic cavity). They are situated cranial to each kidney, and caudal to the pancreatic tissue, with the right testis slightly cranial to the left, and are oval in shape. The testes enlarge during the breeding season, often reaching two to three times their quiescent state. Close to the testes lie the adrenal glands. Each testis has a solitary vas deferens leading down to the urodeum portion of the cloaca, where seminal fluids from the reproductive sexual segment of the kidneys are added.
The male snake also has paired penises, known as hemipenes, in its tail. At rest they are like two inverted sacs either side of the midline and lie ventral to two other small invaginations in the tail which form the anal glands. When a hemipene’s lining becomes engorged with blood, it everts, forming a finger-like protrusion through the vent. As with the domestic cat, the hemipenes are often covered in spines and barbs, and they each have a dorsal groove into which the sperm drops from the cloaca, and so is guided into the female’s cloaca. The hemipenes therefore do not play any part in urination.
Female
The female has paired ovaries, cranial to the respective kidneys, with the right ovary cranial to the left. There are two coiled oviducts starting with the fimbriae opposite each ovary, and moving through the tubular portion of the infundibulum and on into the magnum. From here the tract merges into the isthmus and then the shell gland or uterus before opening into the muscular vagina. This organ ensures that the eggs are laid only when the timing is correct. The vagina empties into the urodeum section of the cloaca. The vascular supply is from the dorsally suspended oviduct mesentery, rather than the caudocranial route employed in mammals.
Most females are stimulated to reproduce in the spring, when the weather warms and the daylight length increases. The tropical boas (such as the boa constrictor) and the Burmese python (Python molurus), however, start breeding when the temperature drops slightly during the cooler portion of the year.
Some species of snake are oviparous (i.e., they lay eggs), others are viviparous (i.e., they bear live young). The latter are, for example, the garter snakes (Thamnophis sirtalis) and the boid family, which have a vestigial egg structure more closely resembling a placenta. Other species make nests, and some species of python will incubate eggs by contracting and relaxing skeletal muscles, so creating warmth.
Sex determination and identification
Snakes are chromosomally dependent for sex determination, as with most mammals. This is in contrast with the Chelonia, Crocodylia and some lizards, in which sex may be temperature dependent.
Sex identification is best made by surgical probing. A fine, sterile, blunt-ended probe is inserted through the vent and advanced just to one side of the midline in a caudal direction. If the snake is a male, then the probe will pass into one of the inverted hemipenes to a depth of 8–16 subcaudal scales. In the female, there are anal glands in this region, and so the probe may be inserted only to a depth of 2–6 subcaudal scales. In some species, such as the boid family, the males possess a paracloacal spur. This is the remnant of the pelvic limb and may be found on either side of body, ventrally, at the level of the cloaca. In very young snakes it may be possible carefully to evert the hemipenes manually, a technique known as ‘popping’.
Skin
The outer epidermal layer in snakes is thrown into a series of folds forming scales, which cover the whole surface of the snake. There are different sizes of scale over the body, with smaller, less raised ones covering the head and larger and more raised scales over the main portion of the body. Some species have scales with ridges on their surface to add greater grip; other species have smooth scales. In some snakes, such as the sea snakes, the skin is very loose fitting, and apparently has few elastic fibres. Other snakes have elastic skin which relatively quickly returns to its normal shape. The reptile skin has little or no skin glands. Its outer layer, or stratum corneum, is heavily keratinised, and composed of three layers of dead cells filled with keratin. These cells become progressively more flattened as they approach the surface. On the ventral surface of the snake there is a single row of scales which span the width of the snake, and are known as the ventral scutes or gastropeges. The caudal edge of each overlaps the cranial edge of the following scale (Figure 17.4).
Figure 17.4 The vent of a rat snake showing the division of the ventral scales from single scutes cranial to the vent to paired scales caudally.

Ecdysis
Ecdysis is the regular shedding of the entire skin. Other reptiles also shed their skin, but the Chelonia and Crocodylia shed individual scutes, and the lizards shed in patches. Only the snakes shed all of their skin (including the clear, fused eyelids, or spectacle) in one go. The stimulus can be dependent on time of year, health status and age of the snake and the process is partly controlled by the thyroid gland.
First, the new layer of skin is formed deep to the old one. Once it is complete, the snake secretes a proteinaceous lymph fluid between the new layer of skin and the old one. At this time the snake will become dull in colour, and often exhibits blueing of the eyes. The fluid forces the outer layer of old skin to separate away from the new, and often contains enzymes to help in this process. Once separation has been achieved the fluid is reabsorbed and the snake’s eyes may be seen returning to normal. A few days later the snake will shed the old skin. It starts the process by rubbing the corners of its mouth on some abrasive surface. The shedding proceeds with the head skin first and the snake then rolls the old skin back until the tail is the last to emerge.
In a healthy snake all of the skin should come away at once. If the skin does not shed the condition is known as dysecdysis. There can be many reasons for this. Disease, dehydration (which causes too little fluid to be produced), scars on the skin surface or lack of an abrasive surface upon which to remove the skin can all be factors. Regular bathing and soft but abrasive damp surfaces may be needed to aid shedding, and any underlying disease should be attended to.
Special cutaneous adaptations in snakes
There are one or two special structures associated with the skin of snakes. These include the lateral spurs of the Boidae, which have been mentioned earlier. The male possesses larger spurs than the female.
Snakes do not have mobile eyelids. Instead, the eyelids have become fused together and transparent, forming the so-called ‘spectacle’.
Many snakes also have special sense organs on the head. The older snake families such as the Boidae have labial pits – a series of depressions running along the dorsal border of the upper jaw. These function as rudimentary heat sensors. In the more evolutionarily advanced species, such as the pit vipers, the heat sensing organs can actually focus on their prey, and are composed of bilateral, forward-facing pits midway between the nares and the eyes. They are supplied by branches of the trigeminal nerves, and, in the case of pit vipers, may be sensitive enough to detect changes of heat as small as 0.002°C!
Snakes do not possess an external eardrum or middle ear. They can, however, hear airborne sounds and can of course detect ground tremors.
LIZARDS
Musculoskeletal system
Lizards have a musculoskeletal system more familiar to those used to dealing with mammalian forms. They possess, in the majority, four limbs, an axial skeleton and much of the anatomical layout of small mammals. There are some exceptions, one being the slow worm, a native of mainland Britain and northern Europe which resembles a snake, having no obvious external limbs. It is actually, however, a highly evolved lizard with rudimentary limbs.
The skull is more rigid than its snake counterpart, having less mobile jaws. There are four rows of teeth, one to each jaw. These are peg-like in shape and are continually replaced in lizards except for the Agamidae and Chamaeleonidae. There are no fang teeth in lizards, but the beaded lizard (Heloderma horribilis) and the Gila monster (Heloderma suspectum) have hollow teeth which allow the venom from sublingual venom glands to ooze through them into the prey when they bite them. These two species are therefore currently classified as dangerous wild animals under the Dangerous Wild Animals Act 1976 (as amended), in the United Kingdom requiring a special licence to keep them in captivity outside of a zoological collection.
The skull articulates with the atlantal cervical vertebra via a single occipital condyle. The thoracic vertebrae and lumbar vertebrae generally have paired ribs on either side. The coccygeal vertebrae possess ventral haemal arches, between which it is possible to access the ventral tail vein for venipuncture.
In many lizards, the tail possesses fracture planes which allow the tail to break off during escape from a predator. These fracture planes occur in the mid to caudal portions of the tail, but not proximally, where vital structures such as the male reproductive organs and fat pads are stored. Only certain species exhibit this tail autotomy. This includes most of the Iguanidae, but does not include the Agamidae, monitor lizards and true chameleons. When the tail is regrown in these species, the coccygeal vertebrae are not replaced, instead a cartilaginous rod of tissue forms the rigid structure. In addition the rows of scales over the new tail surface are often haphazardly arranged and do not match the size and shape of the rest of the tail (Figure 17.5).
Figure 17.5 Regrowth of the tail is possible in many species of lizard, but the scales which regrow are arranged haphazardly, and the vertebrae lost are replaced by a rod of cartilage.

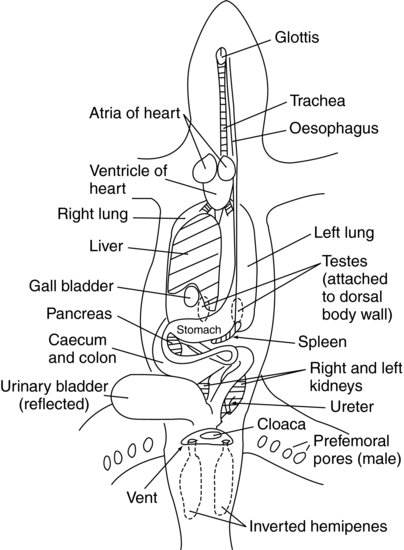
Respiratory system (Figure 17.6)
Upper respiratory system
Lizards have paired nostrils situated rostrally on the maxilla. To the side, or just inside the nares, particularly in iguanids, there is often situated a pair (one on each side) of salt-secreting glands. These are responsible for excreting excess sodium as sodium chloride so helping to conserve water. The sodium chloride may be seen as a white crystalline deposit around the nostrils which is often sneezed out by the lizard. The nostrils enter into the rostral part of the oral cavity, there being no hard palate.
The entrance to the trachea is guarded by a rudimentary larynx which often lacks an epiglottis and vocal folds. Some species, such as the Geckonidae, do possess vocal folds and are capable of producing a variety of sounds. In most species, the trachea is supported by incomplete cartilaginous C-shaped rings similar to those of the cat and dog.
Lower respiratory system
The trachea bifurcates into two main bronchi in the cranial thorax to supply two lungs. In more primitive lizards, the lungs are sac-like structures with large bulla-like divisions and alveoli. In the more advanced lizard species, the lungs are more like the mammalian sponge-like system, with finer divisions and more structured alveolar systems. Lizards will often overinflate their lungs in an attempt to make themselves look bigger, when threatened.
There is no diaphragm in any lizard species, so there is no clear distinction between the thorax and abdomen, rather there is a common body cavity, known as the coelom, as in snakes and birds.
Respiration is thought to be stimulated by falling partial pressures of oxygen in the blood stream. The act of inspiration is due to the mechanical contraction of the intercostal muscles causing an upwards and outwards movement of the rib cage. This is aided by the elastic tissues that are present within the lung structures themselves. Expiration is by contraction of the abdominal and intercostal muscles, and by the elastic recoil of the lung tissue.
Digestive system
Oral cavity
The majority of lizards have a large, fleshy tongue which is frequently mobile. In some species however, such as the chameleons, the tongue has become specialised. It lies coiled in the lower jaw and can be projected out at a flying insect or other potential prey item. The green iguana has a more traditional fleshy tongue, which has a much darker tip. This is not to be confused with pathological changes. A vomeronasal organ is present.
Stomach, associated organs and intestine
The stomach is a simple sac-like structure in most species. The glands are combined hydrochloric-acid- and pepsin-secreting glands lining the walls. There are also separate mucus-secreting glands for lubrication.
The small intestine is better developed in more carnivorous species such as the monitor lizards and the insectivorous water dragons. In herbivorous species it is relatively short. It is poorly divided into jejunum and ileal structures. In a few, mainly herbivorous, species, at the junction between the small and large intestines lies the caecum. The liver is roughly bilobed in structure and is situated ventral to the stomach and lungs. There is usually a gall bladder, with the primary bile pigment being biliverdin, as with birds, rather than the bilirubin of mammals.
The large intestine is more highly developed in herbivorous species than in carnivorous or omnivorous species. Examples include the green iguana and the chuckwalla. These have a large intestine which is often sacculated and divided into many chambers by leaf-like membranes. These increase the intestinal surface area so that microbes, upon which these species depend for vegetation digestion, may colonise it.
The large intestine then empties into the coprodeum portion of the cloaca. The cloaca itself is then continued, as with birds and snakes by the other two segments, the urodeum which receives the urogenital openings, and the proctodeum which is the last chamber before waste exits the cloaca through the vent.
Urinary system
The kidneys are paired and often bean-shaped organs. Their position is variable depending on the species. In some, such as the green iguana, they are both situated in the pelvis, attached to the dorsal body wall (see Figure 17.6). Other species, such as chameleons, have longer kidney structures which extend cranially into the coelomic cavity. As with snakes, the males of some species have a specially developed caudal portion of the kidneys known as the ‘sexual segment’, which enlarges during the breeding season and contributes to the production of seminal fluid. The kidneys empty into the ureters which empty into the urodeum portion of the cloaca.
Many lizards have a bladder. This is not, however, like the sterile bladder of mammals, as it is not connected directly to the ureters. Instead, it is joined to the cloaca, and so urine has to enter the cloaca, before entering the bladder. There is some evidence that the bladder is able to absorb some fluid from its contents, or it may function as a fluid storage chamber, flushing its contents back into the caudal large intestine for further fluid absorption.
Renal physiology
The renal physiology is similar to that already described for snakes. The main differences lie in the variable presence of the urinary bladder, which may have some water reabsorption capabilities.
Cardiovascular system
Heart
The lizard heart is very similar to the snake model, with paired atria and a single common ventricle which nevertheless functions as two. The majority of the deoxygenated blood is channelled to the pulmonary arteries and the oxygenated blood enters the paired aortas.
Blood vessels
The two aortas fuse dorsally, after giving off paired carotid trunks, to form the abdominal aorta. Lizards also possess a hepatoportal venous supply and a renal portal system, hence, as with birds and snakes, intravenous injection into the caudal half of the lizard of medications which are excreted through the renal tubules, could result in their failure to reach the rest of the lizard’s body. It could also increase the toxicity of substances known to be renally toxic if given by this route.
Lizards, like snakes, also possess a large ventral abdominal vein, which returns blood from the tail area and passes just beneath the body wall, ventrally and in the midline. This must be avoided when performing abdominal surgery. This vessel can be used, carefully, for venipuncture for blood sampling in lizards, although the preferred vessel is the ventral tail vein. For intravenous use, the cephalic vein may be accessed on the cranial aspect of the antebrachium, via a cut-down procedure, in the larger species.
Lymphatic system
The lymphatic system is similar to that of snakes, with no discrete lymph nodes.
Reproductive system
Male
The paired testes are situated cranial to the respective kidneys in those species which have abdominally positioned kidneys (Figure 17.6). In those where the kidneys are more pelvic in position, the testes are located just caudal to the end of the lungs and liver, in the middle part of the coelomic cavity. They are supplied by several arteries each and drained by several veins. Both are very tightly adhered to their vascular supply, the left testicle being separated from the left renal vein (into which the left testicular veins drain) by the left adrenal gland. The right testicle is tightly attached to its right renal vein, which separates it from the right adrenal gland. This positioning, so close to such vital structures, makes castrating aggressive lizards a difficult operation. The testes enlarge during the breeding season and regress out of it. Each testis drains into a vas deferens which has a tightly coiled course over the ventral surface of the respective kidney before emptying into the urodeum portion of the cloaca. Some species, such as the Chameleonidae, have a pronounced epididymis extending caudally from each testicle.
The male lizard has paired penises, as with the snake, known as hemipenes. These lie in the base of the tail structure, either side of midline, and function as with the snake family. At rest they are inverted sacs in the tail base. During copulation, one will engorge with blood and evert itself, creating a groove along its dorsal surface. Into this groove sperm and spermatic fluid will drop from the cloaca, and the hemipene will guide this into the female lizard’s cloaca.
Female
The female lizard has much the same anatomy as that described for the snake.
Egg-producing physiology
Reproductive physiology in the female lizard is broadly similar to that of the avian patient. Some species, such as some of the Chameleonidae, are ovoviviparous. That is, they produce live young instead of laying eggs, although the eggs are produced internally. Some species are viviparous, in which a form of placenta or thin-walled egg structure allows the foetus to develop and live young are produced. Many other species are oviparous: that is, they lay eggs. One or two species are parthenogenic: that is, the females produce entire females with no need for a male lizard – some species of Lacerta and Hemidactylus (geckos) are capable of this.
Reptile eggs are generally soft shelled and more leathery than those of their avian cousins. Sexual maturity varies according to the species, green iguanas, for example, reaching it at 2–3 years.
Sex determination and identification
Sex determination is largely dependent on chromosomes. However, geckos as a family are temperature dependent, with 99% of eggs incubated between 26.7–29.4°C being female whereas if the temperature was greater than 32.2°C, 90% of the offspring would be male.
Sex may be identified by surgical probing as mentioned above. This is often the only method available for some species such as the beaded lizard, some monitors and the Gila monster. However, in most other species there are external physical differences. These include the prominent pre-femoral pores of males that are seen on the caudoventral aspect of the thigh of iguanids
(see Figure 17.6). Some male lizards have a series of pre-anal pores just cranial to the vent. Males have wider tail bases than the females to house the large hemipenes (Figure 17.7). Some males have greater ornamentation (Figure 17.8). Male green iguanas and plumed basilisks have larger crests, male Jackson’s chameleons have horns, male water dragons have larger crest spines and many male geckos have a wider vent size and hemipenal bulge.
Figure 17.7 Eversion of a hemipene in a male leopard gecko to remove a hemipenal plug (accumulation of dried secretion).

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


