Chapter 16 Visceral Afferent Systems
GENERAL VISCERAL AFFERENT SYSTEM
The visceral afferent systems consist of neurons whose dendritic zones are located primarily in the viscera of the body as opposed to the somatic afferent system, which innervates primarily the surface of the body. The general visceral afferent (GVA) system consists of neurons whose cell bodies are located in all of the spinal ganglia and in many of the cranial ganglia. Many of the tissues that are innervated are derived from embryonic splanchnopleura. These GVA neurons are concerned with body temperature, blood pressure, gas concentration and pressure, and movement of viscera. The special visceral afferent (SVA) system consists of neurons restricted in their location to specific cranial nerves that function in taste and smell.
ANATOMY
Receptor: Peripheral Nerve
The receptors of the GVA system are located throughout the viscera of the body, where they are stimulated by a number of different modalities. Stretch, distention, and pressure on or in a viscus are the most common modalities. Some receptors are sensitive to chemical changes in the environment. Free endings and a variety of encapsulated endings are found at the dendritic zones of the neurons in this system.7,10,17,19 The axons course over the peripheral nerves that are most readily available to the viscus. In the head, they are the facial nerve to the middle ear and blood vessels of the head; the glossopharyngeal nerve to the caudal tongue, pharynx, carotid body, and carotid sinus; and the vagus nerve to the pharynx and larynx. In the thoracic and abdominal cavities, they are the vagus nerve and the peripheral branches of the sympathetic trunk that include the splanchnic nerves in the abdominal cavity. Here in the body cavities, the GVA axons course toward the central nervous system (CNS) in nerves that contain neurons of the parasympathetic and sympathetic portions of the general visceral efferent (GVE) system (Fig. 16-1). GVA axons from peripheral blood vessels course through the peripheral nerves to the segmental spinal nerves.
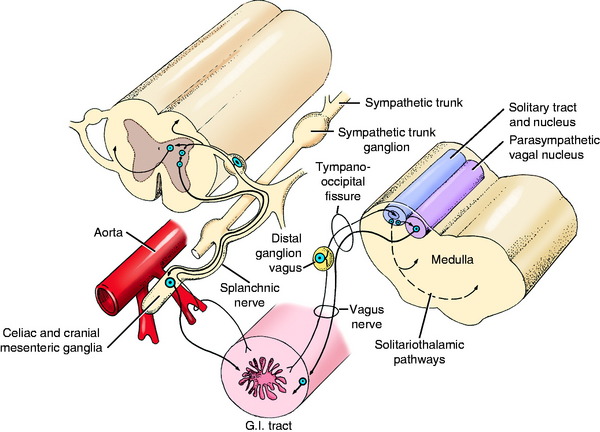
The cell bodies of these GVA neurons are located in the geniculate ganglion of the facial nerve, the distal ganglia of the glossopharyngeal and vagus nerves, and the spinal ganglia of the involved spinal nerves.18 Horseradish peroxidase studies of the esophageal innervation of the dog have shown GVA neuronal cell bodies diffusely distributed in the distal ganglia of the glossopharyngeal and vagal nerves and the cervical and thoracic spinal ganglia.
Brainstem and Tracts
The axons continue from the various ganglia into the CNS. Those axons in the facial, glossopharyngeal, and vagal nerves enter the ventrolateral aspect of the medulla and course to a position near the lateral aspect of the fourth ventricle adjacent to the sulcus limitans. There, the axons course rostrally in a column called the solitary tract (see Figs. 16-1, 2-14 and 2-15). This tract is surrounded by neurons that form the nucleus of the solitary tract in which the axons in the tract terminate. This solitary tract and its nucleus develop in the alar plate adjacent to the motor column of the GVE system. Therefore, it is found dorsolateral to the parasympathetic nuclei of the facial, glossopharyngeal, and vagal nerves. The solitary tract stands out in myelin-stained sections as a densely cylindric structure surrounded by the unstained cell bodies of the nucleus. It extends from the level of the facial nucleus (GSE) rostrally to caudal to the obex caudally. Caudally, the nucleus gracilis and medial cuneate nucleus are dorsal to the solitary tract and rostrally, the medial vestibular nucleus is dorsal to it.
The nucleus of the solitary tract participates mostly in reflex activity and projects to neuronal cell bodies in the GVE system directly or indirectly by way of interneurons in the reticular formation that participate in the various metabolic centers that regulate visceral functions. These include respiratory and cardiovascular activity, swallowing, and micturition. The area postrema receives afferents from the solitary nucleus and participates with the medullary vomiting center.12
The area postrema is located on both sides of the caudal medulla at the level of the obex. It is adjacent to the floor of the fourth ventricle, dorsal and slightly lateral to the parasympathetic nucleus of the vagus. It is a unique parenchymal structure that is readily recognized microscopically by its abundance of small capillaries and glial cells adjacent to a flat layer of ependymal cells that line the fourth ventricle. It is easily mistaken for a neuropathologic lesion. The area postrema is one of the circumventricular organs that have in common the absence of a blood-brain barrier and an anatomic position adjacent to the ventricular system. Other circumventricular organs include (1) the subfornical organ on the caudal surface of the columns of the fornix, where they form the rostral borders of the third ventricle; (2) the subcommissural organ that projects ventrally into the rostral mesencephalic aqueduct ventral to the caudal commissure; (3) the pineal gland that projects caudally from the dorsal portion of the third ventricle; and (4) the hypothalamic vascular organ adjacent to the ventral portion of the third ventricle. These sites are thought to be involved in such functions as chemoreception, neurosecretion, and the control of various visceral activities. They play important roles in the communication between the parenchyma and the cerebrospinal fluid and in communication with other organs via blood-borne products.
Some of the axons of the GVA system receptors in the thoracic and abdominal organs also course centrally over the branches of the thoracic sympathetic trunk and the abdominal splanchnic nerves. Here they are accompanied by the GVE neurons, which are coursing toward the organ innervated (see Fig. 16-1). From the sympathetic trunk, each of these GVA axons courses through a ramus communicans to the segmental spinal nerve, where it enters the dorsal root. The neuronal cell body of the GVA neuron is located in the segmental spinal ganglion. The axon continues over the dorsal root, enters the spinal cord at the dorsolateral sulcus, and continues into the dorsal gray column, where it terminates on a neuronal cell body located there.
In addition to these pathways from the dendritic zones in the gastrointestinal organs to the CNS, there are some, possibly many, GVA neurons that reside entirely within the wall of the viscus in an enteric plexus in which intrinsic reflex activity can occur. They are components of what is referred to as the enteric nervous system, which can function autonomously.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


