Chapter 9 General Sensory Systems
General Proprioception and General Somatic Afferent
SENSORY SYSTEMS
A sensory system is characterized by a peripheral afferent neuron with a dendritic zone that is commonly modified to form a receptor organ; an axon that courses into the gray matter of the central nervous system (CNS); a cell body in a ganglion of the peripheral nervous system; and a centrally located relay nucleus and tract that courses primarily to a specific thalamic nucleus that relays to a specific area of the cerebral cortex. Each type of sensory stimulus, or sensation, is known as a modality, a form of energy that is converted by the receptor organ into a neuronal impulse. These modalities include touch, temperature, movement, chemicals, pressure, light, and sound, which inform the CNS of the features of the external and internal environments of the body. In some instances there is a specific structural neuroanatomic pathway for a modality, such as light and sound. Most receptor organs have a low threshold for a certain modality but still can be stimulated by other modalities. This form of energy to which a receptor organ is most sensitive is referred to as the adequate stimulus. Anatomically there are encapsulated and nonencapsulated receptor organs. The dendritic zone of the encapsulated receptor is associated with a well-developed connective tissue capsule of which there are many varieties. These connective tissue modifications provide the structural features necessary for that receptor to be most sensitive to one specific modality or energy form, although the receptor is not limited to that modality. Several forms of energy may excite that receptor. Even without a connective tissue modification, the nonencapsulated receptors exhibit low thresholds of sensitivity to one specific modality. The histologic features do not necessarily restrict the receptor to sensitivity to one specific modality. Nevertheless, they are classified as thermoreceptors, mechanoreceptors, chemoreceptors, and photoreceptors on the basis of their adequate stimulus.
The use of the term pain is badly abused. I (Alexander de Lahunta) was guilty of this in the first two editions of this book, before Dr. Ralph Kitchell of the University of California set me straight. Pain is not a sensory modality. We do not give a patient a pain stimulus. We do not test for pain. We test for what we interpret as a painful response by stimulating the patient with a noxious stimulus. We test for nociception, which is the patient’s perception of this noxious stimulus. Pain is the subjective cerebral response of the patient to the stimulation of a variety of receptors referred to as nociceptors. This group of receptors is nonselective in the form of energy that elicits its maximal response, but the stimulus threshold for these modalities is high. The intensity of the stimulus required to evoke an impulse from these receptors is at a level that is potentially destructive to the tissue. It requires a high-intensity excitation by mechanical (the pressure of tissue forceps), electrical (a cattle prod), or chemical stimuli. These are noxious stimuli, and the behavioral response observed is nociception, or what we interpret to be pain. The latter varies significantly among individual patients in terms of their response to the same noxious stimulus. Consider the different responses you would expect in reaction to moderate compression of the digits by forceps in a high-strung Chihuahua and in a laid-back collie dog.
GENERAL PROPRIOCEPTION
General proprioceptive (GP) neurons constitute a sensory system that detects the state of the position and the movement in muscles and joints.16 Two basic pathways are described for this system. One is the pathway for segmental reflex activity and for transmitting proprioceptive information to the cerebellum. The other is the conscious proprioceptive pathway that involves the transmission of proprioceptive information to the sensory somesthetic cerebral cortex. These pathways are considered separately for the spinal nerves and cranial nerves.
Spinal Nerve General Proprioception
General Proprioceptive Pathways for Reflex Activity and Cerebellar Transmission
Reflex Activity
The GP axons enter the dorsal gray column of the spinal cord segment. Some axons (1a from a neuromuscular spindle) synapse directly on the general somatic efferent (GSE) alpha motor neuron in the ventral gray column to complete a reflex arc. Others (1b from a Golgi tendon organ) indirectly influence an alpha motor neuron and complete the reflex arc by synapsing on an interneuron (Fig. 9-1). The activity of some interneurons influences GSE alpha motor neurons in other spinal cord segments by passing cranially and caudally in the fasciculus proprius of the lateral funiculus. Recall that this pathway comprises the white matter that is immediately adjacent to the gray matter throughout the spinal cord. It is also referred to as the propriospinal fiber system, which connects adjoining and distant spinal cord segments. This provides a means of interrelating neural activity within the spinal cord.
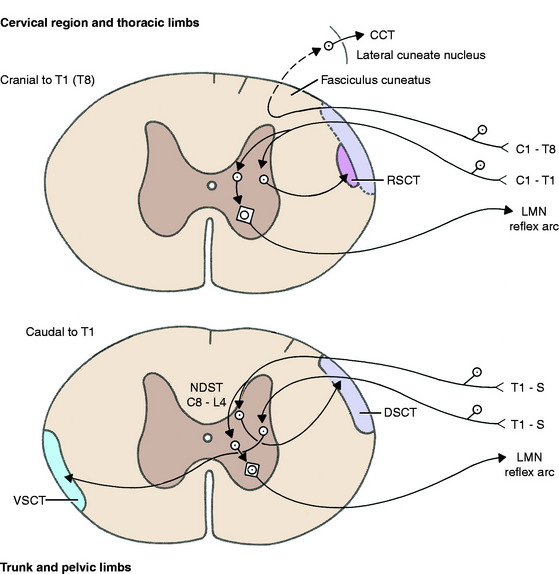
Figure 9-1 GP pathways for reflex action and to the cerebellum-spinocerebellar pathways. CCT, Cuneocerebellar tract; DSCT, dorsal spinocerebellar tract; LMN, lower motor neuron; NDST, nucleus of dorsal spinocerebellar tract—C8-L4 (cat); RSCT, cranial (rostral) spinocerebellar tract; VSCT, ventral spinocerebellar tract.
Dorsal Spinocerebellar Tract
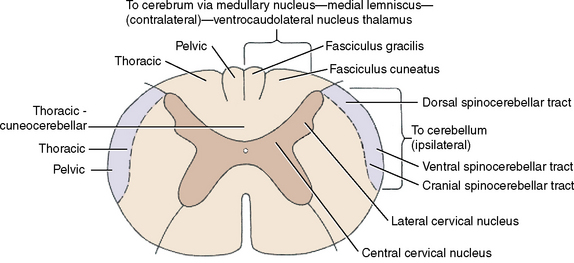
The GP axon enters the dorsal gray column and synapses on a neuronal cell body on the medial aspect of the base of the dorsal gray column. This is in the nucleus thoracicus (nucleus dorsalis, nucleus of the dorsal spinocerebellar tract, Clarke nucleus).21,44 Most of the axons of these cell bodies enter the lateral funiculus of the same side of the spinal cord (ipsilateral) and pass cranially on the surface of the dorsal portion of the lateral funiculus in the dorsal spinocerebellar tract. Here this tract is lateral to the upper motor neuron (UMN) pathways, which includes the lateral corticospinal, rubrospinal, and medullary reticulospinal tracts. The nucleus thoracicus extends from approximately the eighth cervical to the fourth lumbar spinal cord segments in the cat. The GP afferents from the pelvic limbs must course cranially to the cranial lumbar segments to synapse in this nucleus. The dorsal spinocerebellar tract passes cranially through the entire spinal cord to the medulla, where it joins the caudal cerebellar peduncle by way of the superficial arcuate fibers on the surface of the medulla (Fig. 9-4; see also Figure 2.13 through 2.15). It is distributed primarily to the cerebellar cortex of the vermal and paravermal lobules.
Ventral Spinocerebellar Tract
The GP axon enters the dorsal gray column and synapses on cell bodies near its base laterally. These cell bodies form a continuous column from the cranial thoracic spinal cord segments through the lumbar and sacral segments. Most axons of these cell bodies cross to the opposite-side lateral funiculus by way of the ventral white commissure. In the contralateral lateral funiculus, they form the ventral spinocerebellar tract23,32 on the surface of the lateral funiculus ventral to the dorsal spinocerebellar tract. The ventral spinocerebellar tract courses cranially through the entire spinal cord. It continues through the medulla on its lateral side and enters the pons, where it joins the rostral cerebellar peduncle and courses caudally through it into the cerebellum (see Fig. 9-4). Within the cerebellum, most of these axons cross back to the side from which they originated from their cell bodies, and they terminate primarily in the vermal and paravermal lobules.
Cerebellar Transmission from Thoracic Limbs and the Cervical Region
Cuneocerebellar Tract
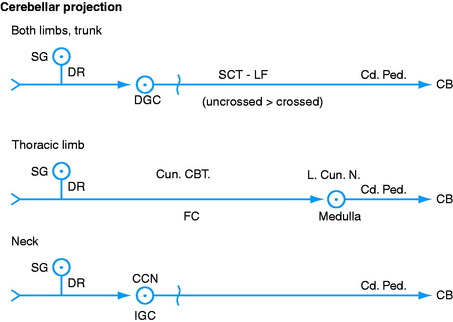
This cuneocerebellar tract contains GP axons from the dorsal roots of the spinal nerves from C1 to T8. It provides a GP cerebellar pathway from the thoracic limbs38 similar to the dorsal spinocerebellar tract from the pelvic limbs.31 The GP axons in the dorsal roots enter the spinal cord along the dorsolateral sulcus and continue dorsal to the dorsal gray column to enter the lateral portion of the dorsal funiculus without synapse. This is the fasciculus cuneatus. The GP axons pass cranially in the fasciculus cuneatus to the caudal medulla, where they terminate by forming synapses with neuronal cell bodies in the lateral cuneate nucleus (see Figs. 9-1 through 9-4; see also Figs. 2-14 and 2-15). This nucleus is located dorsally in the medulla, dorsolateral to the parasympathetic nucleus of the vagus nerve. It is ventromedial to the caudal cerebellar peduncle, rostral to the level of the obex, and caudal to the caudal vestibular nucleus. Axons of the neuronal cell bodies in the lateral cuneate nucleus enter the adjacent caudal cerebellar peduncle and pass into the cerebellum.
Cranial (Rostral) Spinocerebellar Tract
For this thoracic limb GP pathway, the GP axons in the dorsal roots associated with the cervical intumescence enter the dorsal gray column and synapse on neuronal cell bodies near its base in the centrobasilar nucleus.42 The axons of these cell bodies enter the ipsilateral lateral funiculus and course cranially, medial to the ventral spinocerebellar tract (see Fig. 9-2). They continue into the medulla and pons, where they enter the cerebellum through both the caudal and the rostral cerebellar peduncles.
Cervicospinocerebellar Pathway
The central cervical nucleus is located in the intermediate gray column of the first four cervical spinal cord segments.15,28,37 It receives GP axons in dorsal roots from cervical spinal ganglion neuronal cell bodies that are thought to be concerned primarily with general proprioception from the neck (see Fig. 9-3). The axons from the neuronal cell bodies in this nucleus crossto the contralateral lateral funiculus. They continue cranially into the medulla and enter the cerebellum through the caudal cerebellar peduncle.
General Proprioceptive Pathways to the Somesthetic Cortex for Conscious Perception
Fasciculus Gracilis and Fasciculus Cuneatus
These fasciculi located in the dorsal funiculus contain axons that conduct impulses from receptors sensitive to a number of modalities, some of which are concerned with general proprioception. These GP axons are in the dorsal roots of all the spinal nerves. Each dorsal root enters the spinal cord at the dorsolateral sulcus. The GP axons concerned with conscious proprioception pass dorsal to the dorsal gray column and without synapsing enter the dorsal funiculus and course cranially. The GP axons from the pelvic limbs and the trunk caudal to about the level of the sixth thoracic spinal cord segment course cranially in the medial portion of this funiculus in the fasciculus gracilis (see Figs. 9-5 and 9-6; see also Figs. 9-2 and 9-4).36 Cranial to this level, the GP axons are situated more laterally and comprise the fasciculus cuneatus (see Fig. 2-17). The dorsal funiculus is organized somatotopically so that the GP axons from the more caudal levels are positioned in the dorsal funiculus more medially. At progressively more cranial levels, the GP axons are contributed to a more lateral aspect of this funiculus. Thus the cervical GP axons are the most lateral in the funiculus. As these GP axons course cranially, many leave this pathway to terminate in the adjacent spinal cord gray matter.
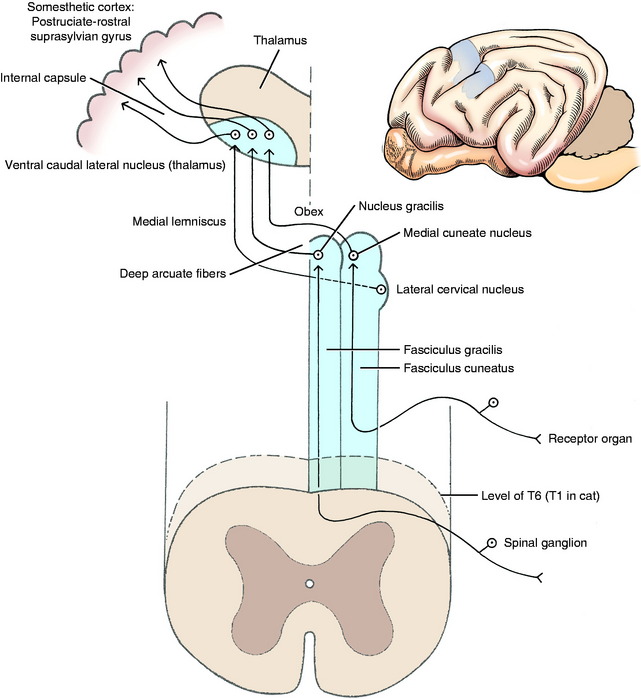
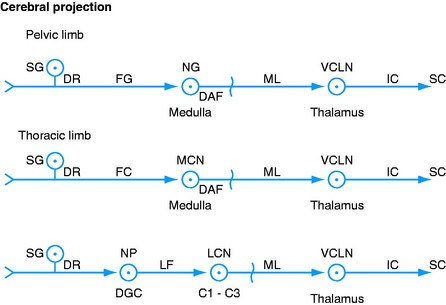
Nucleus Gracilis and Medial Cuneate Nucleus
The GP axons that reach the caudal medulla in the fasciculus gracilis terminate by synapsing on neuronal cell bodies in the nucleus gracilis (see Figs. 9-4 through 9-6).26,39 This nucleus is located in a dorsal position, where it begins in the most cranial portion of the fasciculus gracilis. This occurs just caudal to the level of the obex and at the level of the pyramidal decussation. It is adjacent to the dorsal median sulcus and extends rostrally to the level of the obex, where it is dorsal to the parasympathetic nucleus of the vagus nerve and medial to the medial cuneate nucleus (see Figs. 2-16 and 2-17).
The GP axons concerned with conscious proprioception that reach the caudal medulla in the fasciculus cuneatus terminate by synapsing on neuronal cell bodies in the medial cuneate nucleus (see Fig. 9-5).27,39 This nucleus is dorsal in the caudal medulla just lateral to nucleus gracilis. It is larger than nucleus gracilis and extends rostrally beyond the level of the obex, where it is medial to the caudal portion of the lateral cuneate nucleus (see Figs. 2-15 and 2-16). Remember that the lateral cuneate nucleus is in the cuneocerebellar pathway.
Medial Lemniscus and Ventral Caudal Lateral Thalamic Nucleus
Axons from the neuronal cell bodies in nucleus gracilis and the medial cuneate nucleus course ventrally and transversely through the medulla as the deep arcuate fibers to the opposite side of the median plane (see Figs. 2-14 and 2-15). There they form the medial lemniscus dorsal to the contralateral pyramid and ventromedial to the olivary nucleus (see Figs. 9-5 and 9-6). The medial lemniscus, oriented in a dorsal plane, courses rostrally through the medulla dorsal to the pyramid (see Figure 2.13 through 2.15). As it passes through the dorsal part of the trapezoid body, it is medial to the dorsal nucleus of the trapezoid body (see Figs. 2-11 and 2-12). In the pons, it is located dorsal to the longitudinal fibers of the pons (see Figs. 2-9 and 2-10). In the mesencephalon, it is ventral in the caudal tegmentum and dorsal to the substantia nigra, and it shifts laterally as it course through the tegmentum of the rostral mesencephalon and into the caudal diencephalon (see Figure 2.5 through 2.8). The medial lemniscus terminates by synapsing on neuronal cell bodies in the ventral caudal lateral nucleus of the thalamus. This is a specific projection nucleus of the thalamus for sensory systems that enter the spinal cord over the dorsal roots of spinal nerves. It is a poorly defined nucleus located ventrally in the caudal thalamus dorsal to its external medullary lamina. The axons of the neuronal cell bodies in this nucleus join the thalamocortical fibers and enter the internal capsule and centrum semiovale to be distributed to the somesthetic cortex of the cerebral hemisphere.
The somesthetic cortex is described classically as being located in the parietal lobe of the cerebrum, caudal to the cruciate sulcus.2,25 In the dog, it overlaps with the motor cortex because it is located in the caudal part of the postcruciate gyrus and the rostral suprasylvian gyrus. There is a somatotopic organization of the medial lemniscus, the ventral caudal lateral nucleus of the thalamus, and the somesthetic cortex (see Figs. 9-5 and 9-6). All sensory systems project to localized regions of the cerebral cortex called primary sensory areas. Five of these are well established: auditory, visual, olfactory, gustatory, and somesthetic. The somatotopic organization of the somesthetic cerebral cortex reflects the density of receptor organs in the various regions of the body. For example, the prehensile organs of the animal (the nose and lips of the pig and horse, the lips and tongue of the dog, the forepaws of the raccoon and cat, the hand of the primate) have an abundance of receptor organs for this function and a correspondingly large area of representation in the somesthetic cortex.1,2
Lateral Cervical Nucleus
The lateral cervical nucleus projects from the lateral side of the dorsal gray column in the first two cervical spinal cord segments. Primarily, it receives axons from the spinocervical tract that is located in the ipsilateral lateral funiculus.6,9-11,47,51 The neuronal cell bodies of these spinocervical tract axons are located in nucleus proprius, which is in the middle portion of the ipsilateral dorsal gray column that extends the entire length of the spinal cord. GP axons in all the dorsal roots have access to these neurons in the dorsal gray column.
The neurons in the lateral cervical nucleus serve as the third set of neurons in this spinocervicothalamic pathway.22 The axons of the neuronal cell bodies in this lateral cervical nucleus course cranially and cross to the opposite side in the caudal medulla, where they join the medial lemniscus. Their course in the medial lemniscus to the ventral caudal lateral thalamic nucleus and from there to the somesthetic cortex is similar to that of the deep arcuate fibers that enter the medial lemniscus. The lateral cervical nucleus is somatotopically organized, with pelvic limb afferents synapsing in the dorsolateral aspect of the nucleus and thoracic limb afferents synapsing in the ventromedial aspect. In the dog and cat this nucleus has a major role in the conscious projection of tactile sensation.
Nucleus Z
Once again, the student and clinician should appreciate the complexity that has been introduced here but should also understand the following clinical observations:
Cranial Nerve General Proprioception
Reflex Activity
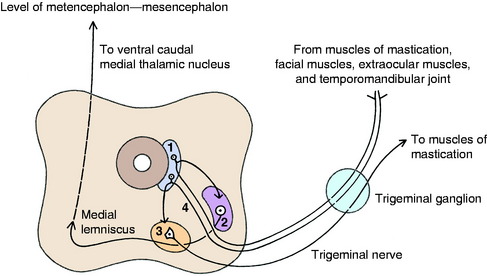
For reflex function, the axons may pass from the mesencephalic nucleus directly to the adjacent GSE nuclei of cranial nerves to synapse on an alpha motor neuron (Fig. 9-7). Other mesencephalic nucleus axons may synapse in the pontine sensory nucleus of the trigeminal nerve which, in turn, may synapse on a GSE neuron in a cranial nerve nucleus.
Conscious Pathway
GP axons from neuronal cell bodies in the mesencephalic nucleus of the trigeminal nerve enter the adjacent pontine sensory nucleus of the trigeminal nerve where they synapse on neuronal cell bodies in this nucleus. This pontine sensory nucleus is located in the caudal pons and rostral medulla, where it is positioned between the trigeminal nerve and its GSE motor nucleus. It is ventromedial to the middle and rostral cerebellar peduncles. The axons of these cell bodies in the pontine sensory nucleus cross through the ventral portion of the reticular formation to join the contralateral trigeminal lemniscus (quintothalamic tract), which joins the medial lemniscus and courses rostrally to the caudal thalamus. These axons terminate by synapsing on neuronal cell bodies in the ventral caudal medial nucleus of the thalamus. The thalamic nuclear axons join the thalamocortical fibers and course through the internal capsule, centrum semiovale, and corona radiata to reach the somesthetic cortex. The involvement of the mesencephalic nucleus of the trigeminal nerve in the GP innervation of head muscles may be limited to the muscles of mastication. Retrograde studies using horseradish peroxidase have shown that the afferent neurons that supply the dog’s tongue have no cell bodies in the mesencephalic nucleus. The cell bodies of the intramuscular neurons of both the intrinsic and extrinsic tongue muscles are located in the first cervical spinal ganglia, the distal ganglia of the vagus, and the trigeminal ganglia. Other studies in the sheep and pig have shown that the neuronal cell bodies of the afferent neurons of their extraocular muscles are all in the trigeminal ganglia.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


