, Monika Hassel2 and Maura Grealy3
(1)
Centre of Organismal Studies, University of Heidelberg, Heidelberg, Germany
(2)
Spezielle Zoologie, Universität Marburg FB Biologie, Marburg, Germany
(3)
Pharmacology and Therapeutics, National University of Ireland Galway, Galway, Ireland
3.1 Fertilization
3.1.1 When Does Life Begin?
Beginnings are often difficult to pinpoint, particularly in life cycles, for life is continuous. Life does not tolerate a break (at most a transitory standstill in a stage of quiescence). Life is a continuing process that started some billions of years ago below the level of multicellularity and will end with the death of the last living being. Nevertheless, each individual life that is based on sexual reproduction has two discrete boundaries: fertilization and death.
Sperm cells and egg cells are not capable of starting an independent life on their own. Once released from the testes or ovary, the lifetime of sperm cells and egg cells is restricted to a few minutes, hours or days (in humans: egg cell 12 h, sperm up to a maximum of 3 days). Only the fusion of sperm and egg results in a cell with the potential to survive and to give rise to an independent individual as well as to a new generation. In biological terms a human is Homo sapiens from fertilization until death. No stage of development can be omitted, no non-human stage defined, at least not in biological terms.
3.1.2 Are Insemination and Fertilization the Same?
Maternal and paternal genomes are brought together in the process of fertilization. Fertilization is preceded by insemination, but these two events are often not so clearly distinguished as some terminological purists want. According to their definition, insemination leads to the encounter of sperm cell and egg cell and culminates in the fusion of the sperm cell membrane with the egg cell membrane; fertilization is the fusion of the two haploid genomes to form the diploid genome of the zygote. A distinction between insemination and fertilization is meaningful and necessary in those organisms where the sperm enters an egg cell before the egg nucleus has completed meiosis. This is the case in mammals as well as in the nematode Ascaris. In such cases the haploid sperm nucleus has to wait until the egg nucleus is also haploid, before fertilization proper can take place. Usually, however, all processes starting with the first contact of sperm cell and egg cell and ending with the fusion of the two nuclei, are grouped under the heading ‘fertilization’.
3.1.3 The Egg Cell Attracts Sperm; Attractants Are Manifold and Their Reception Is Based on Mechanisms Known from Photoreceptors or from Smell and Taste Receptors
The best investigated process of fertilization is that of the sea urchin. Fertilization takes place in the free water column (external fertilization). The freshly spawned egg emits an attracting pheromone in order to guide the sperm to its goal. Pheromones are inter-specific scents which are, as a rule, emitted to prepare mating or to distribute warning signals. In their function as attractants for sperm pheromones are also known as gamones. In the case of sea urchins the gamones are peptides. The flagellum of the sperm is densely equipped with molecular receptors for peptides. Activation of the receptor by bound scents causes a hyper-polarization of the electrical membrane potential, that is an increase in the voltage, as it is known from the photoreceptors (cones and rods) in the retina of our eyes. As mediator between the molecular receptors and the ion channels in the membrane of the flagellum the cGMP-dependent signal cascade is interposed.
Mammalian Sperm.
Even more startling are recent findings about the chemotaxis of the mammalian sperm. It is not so surprising that the chemotactic signals are complex and highly efficient. In experiments sperm cells can be attracted by minute traces (picomol = 10−12 M) of the steroid hormone progesterone. Progesterone is released from the cumulus cells which derive from the follicle and envelope the egg after it has been ejected from the ovary. Progesterone opens ion channels and this stimulates sperm to increase swimming activity. Not only that. Other emitted chemical signals appear to contribute to the attractive features of the egg, for instance prostaglandins and the gaseous nitrogen oxide NO. Most amazing of all, mammalian sperm are equipped with receptors that bind and respond to a substance called bourgeonal, an aromatic aldehyde known as scent emitted by the lily of valley. The substance is effective in extremely low concentrations. However, so far bourgeonal could not be detected in the female genital tract. What is the original odorant adapted to this receptor?
Since sperm swim in a watery fluid, not only typical odorants must be taken in consideration but also typical taste substances. Surprisingly receptors for the taste quality umami have been recently identified in the membrane of the sperm flagellum.
The swimming behaviour of the sperm is mediated by the sperm-specific CatSper channel that controls the intracellular Ca2+ concentration. In humans, CatSper is directly activated by progesterone and prostaglandins – female factors that stimulate Ca2+ influx. Other factors including neurotransmitters, chemokines, and odorants also affect sperm function by changing [Ca2+]i. It appears that the CatSper channel complex serves as a polymodal sensor for multiple chemical cues that assist sperm during their voyage across the female genital tract.
Besides gradients of chemical signals a gradient of temperature along the oviduct is also said to provide guidance for the sperm.
3.1.4 The Sperm Acquires Qualification for Fertilization Only When It Has Undergone a Process of Priming, Called Capacitation
Irrespectively of how sperm is guided up the oviduct it has to swim along this route to become capable of fertilization. It is primed on its way through the uterus and oviduct by substances secreted by the female. This effect is called capacitation of the sperm.
In most mammalian species capacitation serves an additional purpose. For some time the sperm cells get caught by the viscous mucus secreted by the mucosa of the oviduct, and are stored there. Only when ovulation takes place modified products of the oviduct allow swimming to re-commence. Sperm now become hyperactive and escape the obstructive mucus, and they do this just when an egg capable of being fertilized has arrived in the oviduct. As stated above in Sect. 3.1.3, chemo-attractants emitted by the egg itself trigger speeding up of the swimming activity of the sperm.
Capacitation is also required in invertebrates and is, for example, in sea urchins accomplished by the slightly alkaline (pH 8.2) sea water.
3.1.5 The Acrosome, a Chemical Drill, Enables Sperm to Penetrate the Egg Envelopes
The contact of the head of the sperm with components (polysaccharides) of the egg envelope induces the acrosomal reaction. The acrosomal vesicle in the head of the sperm cell (Figs. 3.1, 3.2, and 3.3) opens and releases a collection of hydrolytic enzymes, among them many proteases, bundled in a large protein complex known as the proteasome. In mammals the collection of hydrolases includes glycosidases and hyaluronidases. The sperm possesses a chemical drill head and enzymatically lyses a path through the jelly layer and vitelline membrane, driven by its flagellum. In the sea urchin sperm the drill head elongates to a long boring-rod or finger: from the bottom of the opened acrosomal vesicle a finger-like structure, the acrosomal filament, projects through the enzymatically bored channel penetrating the jelly envelope. The stretching out is accomplished through a rapid polymerization of globular G actin molecules to F actin filaments in the evaginating acrosomal finger (Fig. 3.2). In mammals the zona pellucida is locally dissolved (Fig. 3.3).
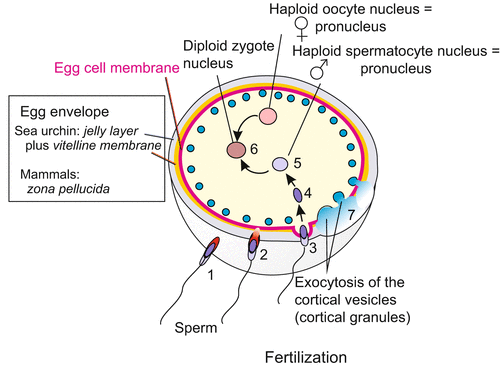

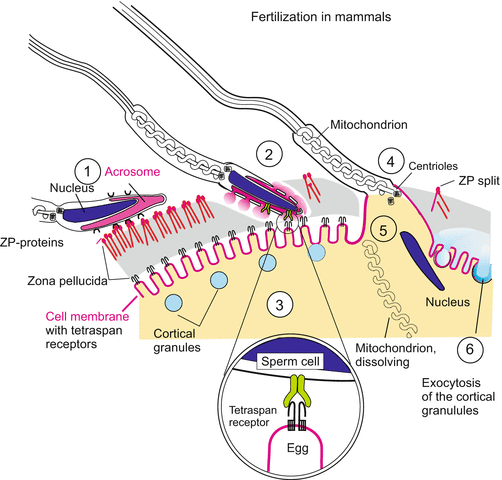
Fig. 3.1
Fertilization in overview. The numbers 1–7 relate to the temporal order of events. (1) Establishment of contact of the sperm with the external egg envelope. (2) Opening of the acrosomal vesicle and drilling through the jelly layer (white) and vitelline membrane (yellow) and contact with the cell membrane (red). (3) Activation of the egg, exocytosis of the cortical granules (blue), lifting off the vitelline membrane (7), uptake of the sperm head into the egg cell (4), release of the sperm nucleus (5) and fusion of the haploid pronuclei to form the diploid nucleus of the zygote
Fig. 3.2
Fertilization in the sea urchin. Contact of the sperm with the jelly coat of the egg induces acrosomal reaction: The acrosomal vesicle opens and release hydrolytic enzymes that begin to digest the envelope at the site of contact. An acrosomal filament is put out and perforates the jelly coat and vitelline envelope; bindin molecules on the surface of the filament establish contact with bindin receptors of the egg cell membrane. Fusion of the cell membranes of the sperm and the egg cell opens a passage through which nucleus of the sperm, together with the centrioles, is drawn into the egg cell. As the cortical vesicles are discharged (cortical exocytosis), the swelling content of the opened vesicles displace and inflate the vitelline envelope. It is now known as the fertilization envelope
Fig. 3.3
Fertilization in a generalized mammal. (1) The sperm head docks with its equatorial zone onto the ZP-protein complex of egg envelope (zona pellucida). (2) Upon contact the acrosomal vesicle is fenestrated and releases a collection of hydrolytic enzymes. These digest a slit into which the rotating spermatozoan moves, propelled by the flagellum. (3) The opening of the acrosomal vesicle exposes a still unidentified ligand (green) that is bound by a complex of tetraspan protein complex in the egg cell membrane. (4) The mutual binding triggers vesiculation of the membranes of the spermatozoon and way is given to the interior compenents of the sperm. The egg cell engulfs (by endocytosis) the nucleus, mitochondrion and centrioles of the sperm. (5) Mitochondria supplied by the sperm are destroyed and degraded in the egg cell while the centrioles are left. (6) By means of exocytosis the cortical granules release enzymes (blue) that degrade the sperm-binding ZP-complex. A cortical reaction similar to that seen in the sea urchin egg and enzymatic linking of zona pellucida proteins provide a hardened fertilization envelope that hinders further sperm from entering the egg cell
3.1.6 Species-Specific Receptors of the Egg Envelope Evaluate the Captured Sperm
Sea urchins. In the free seawater of natural marine environments sperm of several species of sea urchin may contact an egg. It can happen, and is proofed in the laboratory, that sperm of foreign species may attach to the jelly coat and drill their way through the outer envelope of the egg. The jelly coat contains components which trigger the acrosome reaction in sperm from various species. As a final control, sperm and egg check each other out by displaying their identity card: on the outer surface of the acrosomal filament bindin molecules serve as distinguishing signs. Bindins from different species differ in structural details. If the correct bindin is present on the surface of the sperm it will fit onto corresponding bindin receptors in the egg membrane (the extracellular domain of this receptor probably is integrated into the vitelline membrane which encloses the egg cell). This sperm or bindin receptor exists as a disulfide-bonded homotetramer of a transmembrane glycoprotein. Bindin and bindin receptor allow a mutual, species-specific recognition: only sperm of the same species can establish intimate contact with the receptor and thus with the cell membrane of the egg.
The species-specificity of the bindin protein is based on the polymorphism of its gene. Different species of sea urchins possess different variants of the gene which once derived from a common original gene by mutations (point mutations, duplications of certain sequences, deletions). The receptor protein on the egg cell must also have been adjusted to match with the species-specific edition of the bindin signal in a process of co-evolution.
Mammals.
In mammals too the sperm cell must be prepared for its task to penetrate the egg envelopes, and it must be activated on its voyage across the female genital tract at the right moment. On the other side of the sought-after partnership, the egg must be enabled to catch and hold back the sperm. For that purpose glycoproteins are exposed in the zona pellucida of the egg which surrounds the egg cell proper. In the mouse these glycoproteins are known as ZP1, (ZP = zona pellucida), ZP2 and ZP3 (in humans an additional ZP4 has been found). The ZP3 protein has been considered to be the primary sperm-catching molecule but definitive proof is still lacking. Recent evidence indicates that the three-dimensional ZP complex as such, and not a single constituent, mediates sperm adhesion.
All ZP-proteins possess cystein-rich extracellular domains similar to those known from many other extracellular domains. Together the ZP proteins form a fibrillar complex that exists in a form allowing the head of the sperm to couple with it only in the zona pellucida of non-fertilized eggs. After fertilization some components of the complex are enzymatically cleaved and the sperm-binding capacity of the zona pellucida decays.
The counterpart of the ZP-receptor-complex in the envelope of the egg is its ligand in the head of the sperm. In mammals the sperm comes in contact with the egg envelope not with its tip but with its equatorial zone. A particular binding zone extends along the surface of the equator that contains several factors indispensable for fertility. One of these proteins has been designated ESP (Equatorial Segment Protein) which in cooperation with a series of other proteins such as SED1 mediates the binding to the ZP complex of the egg. Binding triggers the acrosomal reaction and further proteins become exposed on the surface of the sperm.
However, the sperm catching proteins of the zona pellucida are not integral proteins of the egg cell membrane, the oolemma, but components of the extracellular envelope. Therefore, a further process must follow that allows the fusion of the sperm cell with the egg cell. Fusion is preceded by vesiculation of the outer acrosomal membrane of the sperm. The ultimate terminal binding partners that mediate fusion of the vesicles with the egg cell membrane, the oolemma, are still unknown at present.
Hypotheses.
Several molecules have been proposed as binding partners on the side of the egg cell, among them a member of the integrin family and fertilin-β, a protease. But subsequent analyses have defeated initial promising reports. A recent hypothesis assigns mediation of sperm-oolemma adhesion to a transmembrane protein of the immuno-superglobulin family (IZUMO) on the side of the sperm head and four-transmembrane proteins, collectively known as tetraspanins (actually CD9 and CD81) on the oolemma. A general property of tetraspanins is their ability to interact with one another and many other surface proteins and, thus, to mediate cell adhesion and fusion. Actual egg-sperm fusion is thought to share similarities with synaptic vesicle fusion. Integrins may play a supporting role via sustenance of the tetraspanin web in the oocyte cortex.
A Preliminary Summary.
In parallel to the events known from sea urchins (and from other ‘fertilization models’ such as ascidians) and supported by laboratory experiments with mouse gametes, the following sequence of events has been proposed: The mammalian sperm first communicates with the ZP-complex of the egg envelope. The ZP-complex attaches onto the sperm head and stimulates exocytosis of the acrosomal vesicle. A collection of hydrolytic enzymes is released, among them proteases bundled together to form a proteasome. The machinery of the proteasome attacks ubiquinated proteins of the egg envelope, the zona pellucida, and a channel leading to the egg cell is enzymatically cleared. Now the acrosomal membrane can establish immediate contact to the oolemma, the egg cell membrane proper. The acrosome exposes a protein, functionally comparable to the bindin in sea urchins, and presumably represented by a member of the highly polymorphic immunoglobulin family. The particular species-specific immunoglobulin serves as an identity card and becomes the ligand that couples the sperm cell to the ultimate receptor in the egg cell membrane. This receptor is represented by, or integrated in, a collection of tetraspan proteins. The ligand-receptor binding enables fusion of the membranes of the sperm cell and the egg cell.
Mutual adhesion triggers a signal transduction cascade similar to that shown in Fig. 3.5 for the sea urchin egg and described in Sect. 3.2. Following membrane fusion, the internal contents of the spermatozoon – its nucleus, mitochondrion and centrioles – are drawn into the egg. The egg’s surface bulges outward into a fertilzation cone, and long microvillar processes erupt from the surface towards the sperm. The processes stretch out and grasp the head of the spermatozoon. Its nucleus, mitochondria and centrioles become engulfed.
3.1.7 As a Rule, Entrance Is Refused for Further Sperm Cells
When the cell membranes of sperm and egg fuse, entry of further sperm should be prevented. The biology of fertilization distinguishes between two mechanisms which both serve to block polyspermy (the entrance of multiple sperm). Both mechanisms are switched on during activation of the egg (Sect. 3.2):
1.
The primary, fast but only transient polyspermy block is thought to be mediated by a conformational change of the three-dimensional structure of the sperm-binding receptors on the egg, which now prevents additional sperm from attaching. This change is induced by a depolarization of the electrical cell membrane potential (decrease of the voltage), triggered by the sperm-egg adhesion.
2.
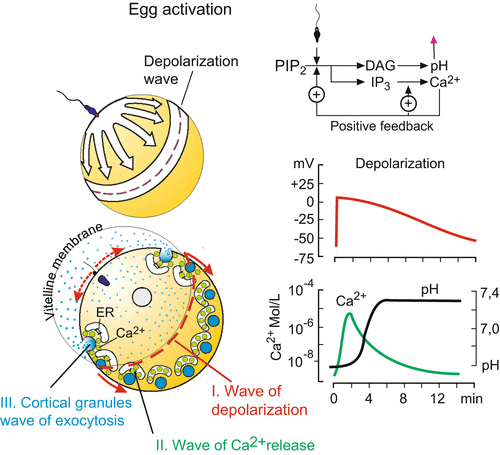
The secondary and permanent block is realized by the rapid inflation of the so called fertilization membrane which suddenly sets apart from the egg surface and lifts off (more details in Sect. 3.2). This inflation can be seen clearly in the eggs of sea urchins and amphibians. In mammals the process of permanent blockage is less conspicuous and brought about by enzymatic degradation of the ZP-complex; thus, no further sperm can dock.
Often, inactivation of the binding molecules is vital. In most animals polyspermy is disastrous and causes early death of the embryo. Amphibian and avian eggs, however, appear to tolerate polyspermy; supernumerary sperm are destroyed within the egg.
3.2 Activation of the Egg
3.2.1 Sleeping Beauty Is, with Awakened With a Kiss
The unfertilized egg sleeps: transcription, protein synthesis, and cell respiration are at or near zero levels. Contact of the acrosomal filament with the vitelline membrane triggers a cascade of dramatic events; they are best studied in sea urchins but occur in the mammalian egg in similar way (Figs. 3.4 and 3.5).