CHAPTER 3 The Role of Wolbachia in Heartworm Infection
Wolbachia are gram-negative bacteria that reside within the body of many filarial nematodes, including D. immitis. From the moment Sironi et al1 discovered that D. immitis harbors Wolbachia, the scientific community realized that a major discovery had been made, one that would likely change the way they looked at filarial disease. Indeed, as gram-negative bacteria, Wolbachia have the potential to play an important role in the pathogenesis and immune response to filarial infection. The immunopathology of filarial disease is extremely complex, and the clinical manifestations of infection are strongly dependent on the type of immune response elicited by the parasite. These bacteria are known to play a fundamental role in worm biology, reproduction, and long-term survival. Less is known about what role they may play in the pathogenesis of heartworm infection. It is likely that the proinflammatory reaction to these bacteria may exacerbate the clinical picture observed in heartworm-infected animals, including cats.
ETIOLOGY
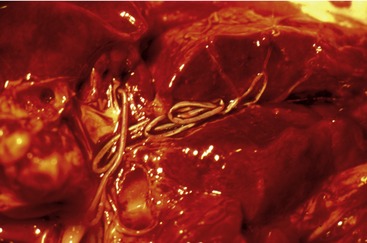
Feline heartworm disease (FHWD) is caused by the filarial nematode D. immitis, which resides in the pulmonary arteries of infected cats (Figure 3-1). Female worms are approximately 15 inches in length, whereas males are shorter. They are less than 5 mm wide. Contrary to what is seen in dogs, naturally infected cats usually harbor less than six adult worms. Female worms release first-stage larvae (microfilariae) into the bloodstream and these are taken up by competent mosquito vectors. After approximately 15 days, these larvae develop into the infective third-stage larvae (L3) and can be inoculated into a new host. Immature worms migrate first through connective tissue and then into the venous circulation, where they are carried to the pulmonary arteries. Worms are fully mature approximately 8 months after cats become infected. There is evidence that only a small portion of infective larvae reach full maturity in infected cats, likely due to a strong immune response against the immature worms. Cats also can eliminate circulating microfilariae and reduce the reproductive capacity of adult female worms.2
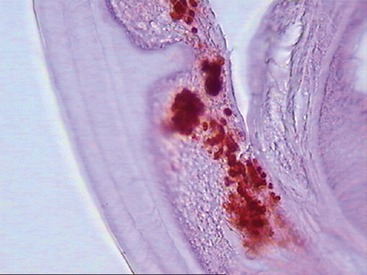
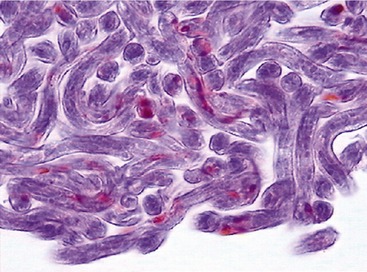
Wolbachia has been identified as an endosymbiont of many arthropods and filarial nematodes. Wolbachia pipientis, the only species thus far identified in the genus, are gram-negative bacteria belonging to the order Rickettsiales. They are closely related to other bacteria belonging to the same group, such as Ehrlichia spp. and Anaplasma spp., as shown in Figure 3-2.3 Electron microscopy, histological examination, and immunohistochemistry have offered a clear description of the distribution of Wolbachia in D. immitis.4 They are found throughout all the stages of the life cycle of the nematode, although they occur in varying proportions between individual worms and different developmental stages. In adult D. immitis, Wolbachia is found predominantly throughout the hypodermal cells of the lateral cords (Figure 3-3). In female worms, Wolbachia also is present in the ovaries, oocytes, and developing embryonic stages within the uteri (Figure 3-4). They have not been demonstrated in the male reproductive system, suggesting that the bacterium is maternally transmitted through the cytoplasm of the egg and not through the sperm. There are several features of the relationship between Wolbachia and filarial worms (including D. immitis) that suggest its symbiotic nature: (1) in those species of filarial worms that have been identified as harboring Wolbachia, all of the individuals are infected (i.e., 100 per cent prevalence); (2) the evolution of the bacteria match that of the filarial worms, as shown by phylogenic studies; (3) the bacteria are transmitted from female to offspring (thus the symbiont guarantees its own future by increasing the fitness of the host that is involved in its transmission); and (4) removal of Wolbachia (antibiotics/radiation) leads to sterility of female worms and eventual death of adult worms.
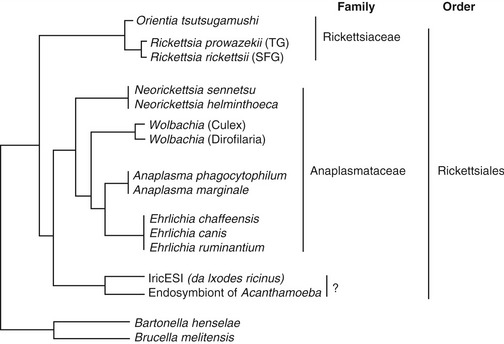
Figure 3-2 The position of Wolbachia within the order Rickettsiales, based on phylogenetic analysis.
EPIDEMIOLOGY
Numerous studies have been carried out to determine the prevalence of exposure to D. immitis infection in cats from different parts of the world. Reliable results are hindered, however, by the difficulty in diagnosing patent infection in cats (see section Diagnosis). Most studies distinguish rates of mature infection (presence of at least one adult worm in the pulmonary artery, usually at necropsy) from immature or transient infection (presence of circulating antibodies against worm antigens by serology). A large-scale study carried out on more than 2000 cats in the United States5 reported an average rate of heartworm antibodies of 12 per cent, with peaks of more than 30 per cent in certain states like California and Florida. In a more recent study in Florida,6 heartworms were identified at necropsy in 4.9 per cent of 640 cats in a shelter, and serological evidence of heartworm infection was present in 17 per cent. Studies from endemic areas in Europe have reported similar findings. We carried out a large study in an endemic region of Italy7 and tested more than 1000 cats for circulating microfilariae, antigen, and antibodies against D. immitis. Results showed that 16 per cent of all tested cats were positive for anti–D. immitis antibodies, with values ranging from 9 to 27 per cent, depending on location. Of these, 3 per cent were positive on antigen testing and 4 per cent were microfilaremic (see section Diagnosis). It is generally accepted that the prevalence of heartworm infection in cats is approximately 5 to 10 per cent of the prevalence in dogs in the same geographical area.
Heartworm-infected cats show evidence of exposure to Wolbachia. Indeed, a varying percentage of both experimentally and naturally infected cats8,9 present circulating antibodies against a major surface protein of Wolbachia (Wolbachia surface protein [WSP]). It is not clear at what point(s) during the infection the bacteria can leave the body of the worm and be released into the blood/tissue of the host. Because all heartworms harbor the bacteria, and because the bacteria are present in all life stages of the worm, Wolbachia could be released by worms during the molting process, when they shed their old cuticle to form a new one, during reproductive activity, when mature female worms release microfilariae into the bloodstream along with debris from the uterus, or following death of the worms (e.g., microfilarial turnover, spontaneous death of adults). As mentioned above, most infective larvae die before they mature to adulthood in cats. Interestingly, it would appear that the death of infective larvae also could cause exposure to Wolbachia in cats. It has been shown that in experimentally infected cats, there are very high levels of anti-WSP antibodies starting at about 2 months postinfection. At this time, many L3 and young L4 larvae will have been eliminated by the host’s immune system, and death of these larvae likely will have allowed the release of the bacteria into the surrounding tissue. These results were confirmed when another group of cats were treated with ivermectin at 2 months postinfection to induce the death of all larvae; antibody titers against Wolbachia were high and persistent.9 Levy et al,10 however, recently carried out a large-scale serological study in cats and did not report a significant correlation between infection status of cats, as determined by serology/necropsy, and the presence of anti-WSP antibodies. Clearly, there is still work to be done in determining how often cats exposed to heartworm infection also are exposed to Wolbachia.
PATHOGENESIS
Although live adult worms in the pulmonary arteries cause a local arteritis, some cats never manifest clinical signs. When signs are evident, they usually develop during two stages of the disease: (1) arrival of heartworms in the pulmonary vasculature and (2) death of adult heartworms. The first stage coincides with the arrival of immature adult worms in the pulmonary arteries and arterioles: approximately 3 to 4 months postinfection. These early signs are due to an acute vascular and parenchymal inflammatory response to the newly arriving worms and the subsequent death of most of these same worms. This initial phase is often misdiagnosed as asthma or allergic bronchitis, but in actuality is part of a syndrome now known as heartworm-associated respiratory disease (HARD).11 Clinical signs associated with this acute phase subside as the worms mature, but demonstrable histopathological lesions are evident even in those cats who clear the infection. Browne et al12 recently reported that cats with serological evidence of exposure to heartworms, including those without adult heartworms in the lungs and heart, have a greater prevalence of pulmonary arterial lesions than heartworm-negative cats without serological evidence of exposure.
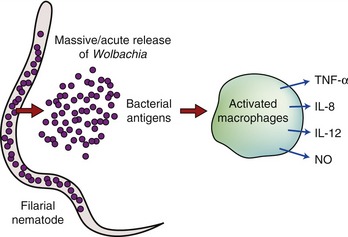
The nature of the worm antigens that cause the intense inflammatory reaction seen in cats is still not known. Immature and adult worms likely possess myriad candidate antigens and Wolbachia most certainly is among them. There is no evidence to date that Wolbachia is able to infect mammalian cells. However, in filarial infected hosts, including cats infected by D. immitis, the release of bacteria following worm death has been shown to be associated with the upregulation of proinflammatory cytokines, neutrophil recruitment, and an increase in specific immunoglobulins.13 Therefore the role of Wolbachia in the host response to filarial infection includes interaction between bacterial molecules and the innate and adaptive immune system.
The innate immune system represents a defense mechanism against molecular structures that are conserved among a wide range of organisms. It consists of the recognition of specific “markers” (pathogen-associated molecular patterns [PAMPs]) that signal the presence of “generic” pathogens. The consequent recognition of these PAMPs by toll-like receptors (TLR) on the surface of antigen-presenting cells leads to the production of reactive oxygen species, proinflammatory cytokines, and to the upregulation of costimulatory molecules that assist in development of an adaptive immune response (Figure 3-5).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


