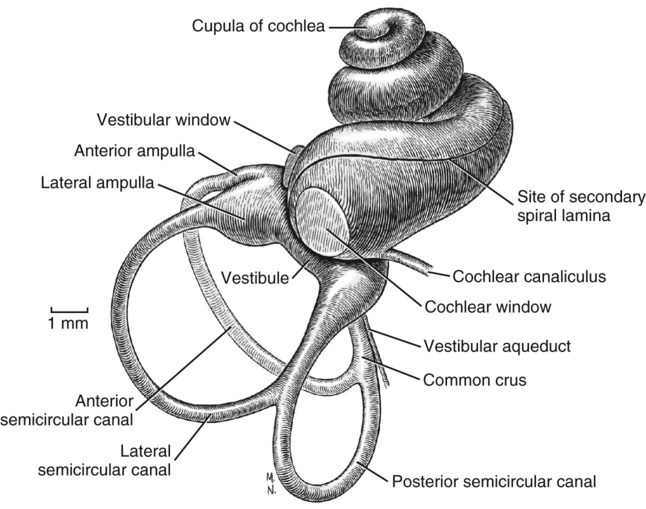
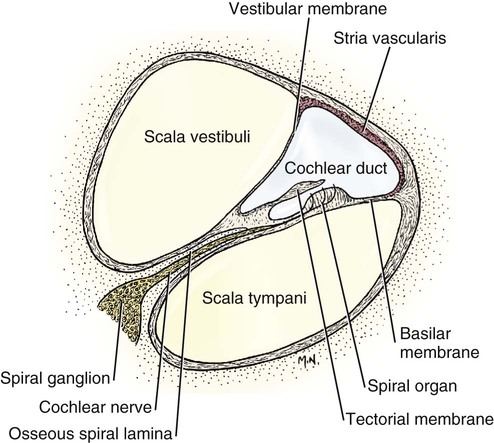
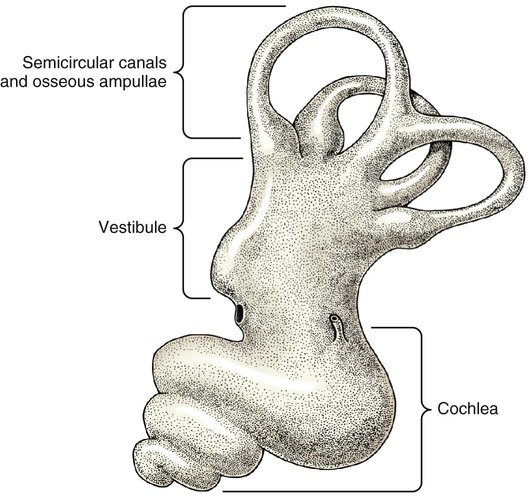
The three major components of the dog ear will be considered here in developmental sequence, which is similar to their phylogenetic evolution: (1) the inner ear, (2) the middle ear, and (3) the external ear (Fig. 20-1). Corresponding developmental modifications occur in the surrounding paraxial mesoderm to provide a supporting capsule for the membranous labyrinth. This fluid-filled (perilymph) ossified structure is the bony labyrinth contained within the developing petrous portion of the temporal bone (Fig. 20-1). There are three semicircular canals, an anterior, a posterior, and a lateral canal (Figs. 20-2 and 20-3). They lie caudal and slightly dorsal to the vestibule. Each canal describes approximately two thirds of a circle in a single plane, and each is approximately at a 90-degree angle to the other two. The segment of the canal that communicates with the vestibule is called the crus. Each canal has two crura that communicate with the vestibule (with the exception of the common crus, to be noted later). One crus of each canal has a dilation, the osseous ampulla (ampullae osseae) near the junction with the vestibule. The lumen diameter of the canals averages roughly 0.5 mm, the ampulla being approximately twice as large. The cochlea is the bony shell that surrounds the cochlear duct in a spiral of three and one-quarter turns around a central hollow core of bone, the modiolus, which contains the cochlear nerve and blood vessels. The cochlea points ventrorostrally and slightly laterally within the promontory of the petrous part of the temporal bone (Fig. 20-4). The osseous spiral lamina that winds around the modiolus, much like the thread of a screw, nearly bisects the lumen of the cochlea into two portions, called the scala tympani and scala vestibuli (Fig. 20-5). The osseous spiral lamina begins within the vestibule and ends at the apex in a free hooklike process, the hamulus. The scala vestibuli communicates with the vestibule, and hence the perilymph within it is acted on by the vibrations of the base of the stapes in the vestibular window. The cochlear window (see Figs. 20-1 and 20-3) is an opening situated near the rostral end of the vestibule by which the scala tympani communicates with the tympanic cavity. A secondary tympanic membrane closes this cochlear window. The membranous cochlear duct, formerly scala media, completes the separation of the two scalae. The scalae communicate at the apex of the modiolus by a small opening, the helicotrema, formed at the free border of the hamulus. The basal turn of the cochlea is approximately 4 mm in diameter and lies close to the medial side of the vestibule. The total height of the cochlea measures approximately 7 mm. Longitudinal modiolar canals and a spiral modiolar canal serve for the distribution of both blood vessels and nerves to the cochlea. The source of perilymph may depend on its location. One source is CSF from the subarachnoid space that gains entrance to the scala tympani of the cochlea via the perilymphatic duct (ductus perilymphaticus) (not part of the membranous labyrinth) in the small cochlear canaliculus (canaliculus cochleae). This small canal courses directly ventrad from a point on the ventral wall of the scala tympani near its origin to communicate with the subarachnoid space (see Fig. 20-1). The other source is as an ultrafiltrate from the cochlear blood vessels in the modiolus into the scala vestibuli. The dendritic zone of the vestibular portion of cranial nerve VIII is in a synaptic relationship with the hair cells of each crista ampullaris and the macula utriculi and macula sacculi. The axons course through the internal acoustic meatus with those of the cochlear nerve. The cell bodies of these bipolar-type sensory neurons are inserted along the course of the axons within the petrous portion of the temporal bone, where they form the vestibular ganglion (Giene and Kuder 1983). The vestibular nerve axons pass to the lateral surface of the rostral medulla where they enter the medulla and terminate in telodendria at one of two sites. The majority terminate in the vestibular nuclei in the medulla and pons. A few course directly into the cerebellum by way of the caudal cerebellar peduncle. The most highly developed and differentiated portion of the membranous labyrinth is the cochlear duct (Figs. 20-5 and 20-6). This duct has a triangular shape with its base, the stria vascularis adjacent to the peripheral wall of the cochlea. A thin vestibular membrane forms the roof of the cochlear duct and a thicker basilar membrane forms the floor of the duct. The spiral organ (organum spirale), formerly organ of Corti, is a collection of hair cells and supporting cells that rests on the basilar membrane. These structures are involved in the transduction and transmission of sound impulses via the cochlear nerve to the brain. Special somatic afferent axons in the cochlear nerve make synaptic contact with these hair cells. Their neuronal cell bodies are centrally located in the attachment of the spiral lamina to the modiolus. The axons leave the inner ear through the internal acoustic meatus accompanied by the vestibular nerve and synapse in the cochlear nuclei on the lateral side of the medulla oblongata. The basilar membrane separates the endolymph of this duct from the perilymph of the scala tympani, which is a part of the cochlea. The thin vestibular membrane separates the endolymph of the cochlear duct from the perilymph in the scala vestibuli of the cochlea. Thus the membranous cochlear duct filled with endolymph is enclosed within the bony cochlea (scala tympani-scala vestibuli), which is filled with perilymph. These fluids have different chemical compositions and are not in open communication with each other. Likewise the membranous semicircular ducts containing endolymph are enclosed within the semicircular canals that contain perilymph. The distinction between the osseous (Fig. 20-7) and the membranous labyrinth (see Fig. 20-6) is sometimes blurred in textbooks by the carefree use of canal for duct and cochlea for cochlear duct. To understand the structural and functional relationships of the labyrinths to each other, the terminology must be kept clear and consistent. Hearing depends on the ability of sound waves in the external gaseous environment reaching the tympanic membrane via a patent external acoustic meatus. Here the oscillations in the air are converted to oscillations in the three auditory ossicles in the tympanic cavity. At the vestibular window the bony oscillations are converted to oscillations of the perilymphatic fluid, which in turn affects the spiral organ by oscillations of the basilar membrane in the cochlear duct. Any interruption of this pathway can result in loss of hearing. Most deafness is related to disorders of the spiral organ. Foss and Flottorp (1974) found that hearing in puppies first occurred as a functional modality at 14 days after birth (on average), which coincides with the opening of the eyelids. Congenital inherited sensorineural deafness is very common in many breeds of dogs and is usually present shortly after birth. Johnsson et al (1973) studied deafness and pathology of the cochlear in Dalmatian dogs. Rouse et al (1984) considered abnormal otoconia and calcification of the labyrinth in deaf Dalmatian dogs. Albinotic and abiotrophic forms are recognized and can be diagnosed in puppies using brainstem auditory response testing (de Lahunta and Glass, 2009.) Shambaugh (1923) injected the vessels of the dog labyrinth and found that the arterial supply was from a single labyrinthine artery that entered through the internal acoustic meatus. In some specimens a small artery came along the cochlear canaliculus and was distributed to a small area of periosteum along the scala tympani. The first branch to be given off from the labyrinthine artery was the anterior vestibular artery, a vessel that supplied the macula of the utriculus and the cristae of the lateral and anterior semicircular ducts. The second branch of the labyrinthine artery supplied the crista of the posterior duct, the posterior crura of the posterior and horizontal canals and ducts and the crus commune. The venous drainage from the labyrinth was collected by two trunks, the larger one leaving along the cochlear canaliculus collected all of the blood from the cochlea as well as most of the blood from the capillaries supplied by the anterior and posterior vestibular arteries. The lesser vein leaving along the vestibular aqueduct drained the remainder of the labyrinth.
The Ear
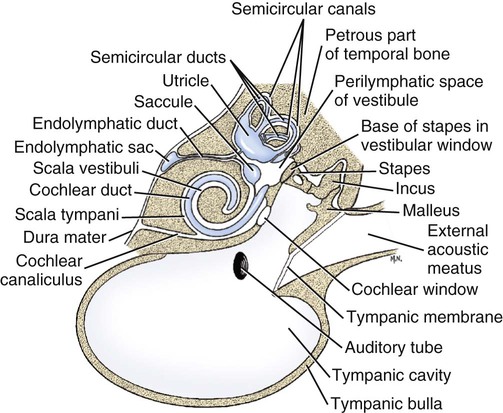
The Inner Ear
Bony Labyrinth
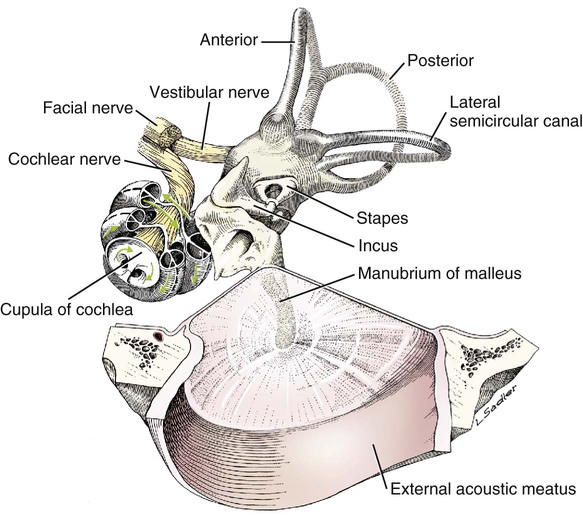
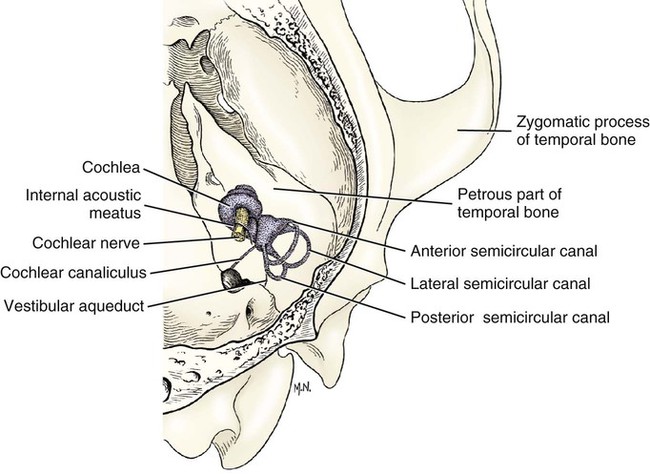
Semicircular Canals
Cochlea
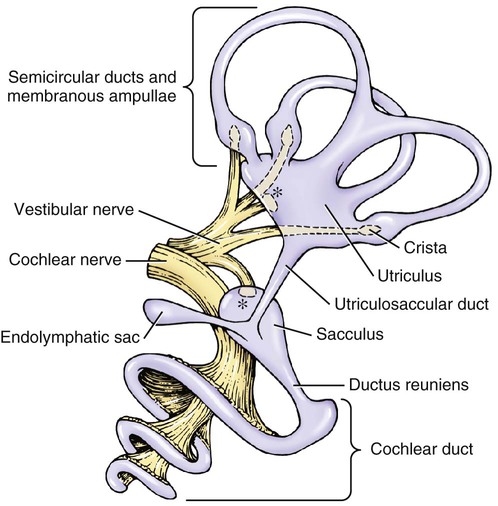
Membranous Labyrinth
Vestibular Nerve
Cochlear Duct-Spiral Organ
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The Ear
Only gold members can continue reading. Log In or Register to continue
