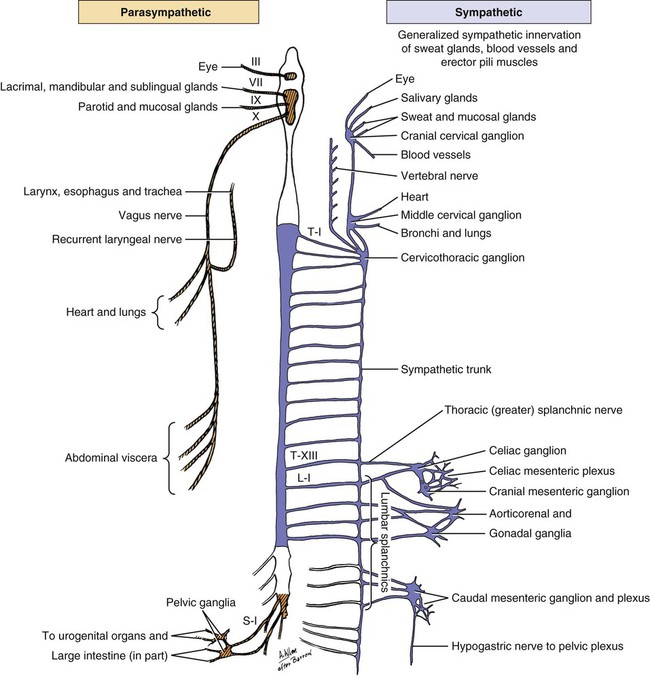
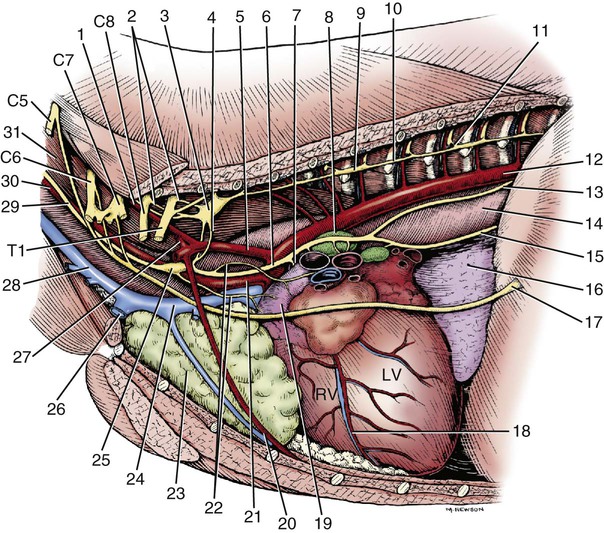
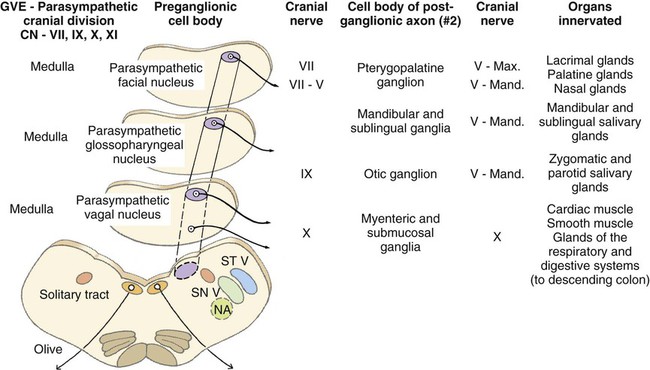
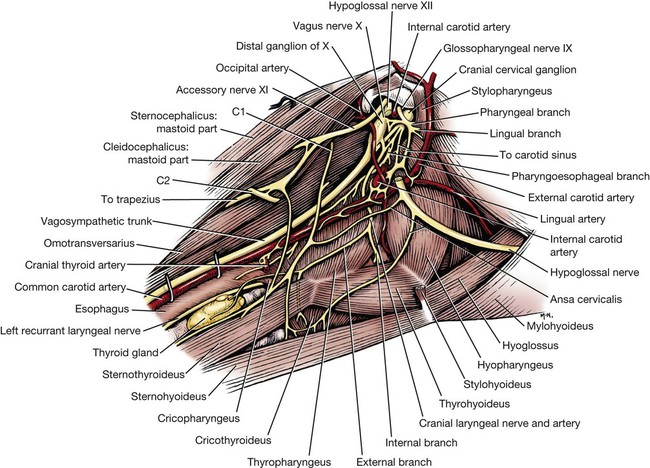
Parasympathetic preganglionic axons leave the brainstem as part of cranial nerves III, VII, IX, and X (Figs. 15-1 and 15-2). The axons in nerves III, VII, and IX are distributed to the head region, whereas the vagus nerve distributes autonomic axons to the cervical, thoracic, and abdominal viscera as far caudally as the left colic flexure. Parasympathetic preganglionic axons also leave the spinal cord as part of the ventral roots of the sacral nerves and become part of the pelvic plexus. It is this brainstem and sacral spinal cord origin of parasympathetic axons that is described by the term craniosacral as a synonym for parasympathetic. Other routes of distribution may exist. For example, a rich and widely distributed cholinergic innervation of cerebral blood vessels has been described in the dog (Amenta et al., 1980). The nerves are restricted mainly to the adventitia, are more prominent in arteries, remain intact after removal of the cranial cervical ganglia, and are therefore assumed to be parasympathetic. Preganglionic neurons lie in the parasympathetic nucleus of the oculomotor nerve (nucleus parasympathici oculomotorii) which is unpaired in the dog and is located on the median plane just ventral to the central gray substance of the rostral mesencephalon (Figs. 15-1 and 15-2). This nucleus is called the nucleus of Edinger-Westphal in humans. The axons run as part of the third cranial nerve through the middle cranial fossa to leave the cranial cavity via the orbital fissure. They leave the oculomotor nerve by a short root and terminate at the ciliary ganglion (ganglion ciliare). Synapse occurs in the ciliary ganglion, and the postganglionic axons leave as the short ciliary nerves. They supply the ciliary muscle, which regulates lens curvature, and the sphincter of the iris, which, when activated, reduces pupillary diameter. Distribution of cholinergic as well as adrenergic innervation in the eye is described (Gwin et al., 1979). An accessory ciliary ganglion has been described in 13 mammalian species, including canines in which it is located approximately 4 mm from the ciliary ganglion on one of the short ciliary nerves (Kuchiiwa et al., 1989). The cell bodies of the preganglionic parasympathetic neurons associated with the facial nerve form two small nuclei in the medulla, the parasympathetic nuclei of the facial and intermediate nerves (Figs. 15-1 and 15-2). The parasympathetic nucleus of the facial nerve (nucleus parasympatheticus n. facialis) is closely associated with the somatic motor facial nucleus (Azuma et al., 1983). The preganglionic axons of these small cell bodies emerge from the medulla in the pars intermedia of the facial nerve, the intermediate nerve (n. intermedius) which immediately follows the facial nerve. Within the facial canal of the petrosal portion of the temporal bone, these preganglionic axons leave the facial nerve in the major petrosal nerve (n. petrosus major) and continue into the orbit in the nerve of the pterygoid canal (n. canalis pterygoidei) (Wakata, 1975) to terminate in synapses in the pterygopalatine ganglion (ganglion pterygopalatinum) located medial to the sphenopalatine nerve on the surface of the medial pterygoid muscle. Postganglionic axonal distribution to the lacrimal gland is not well defined. Apparently, some axons may go directly as small branches, and some may run with the lacrimal and zygomatic nerves. Other postganglionic axons go to glands and smooth muscle of the nasal and oral cavities via branches of the fifth cranial nerve (Jung et al., 1926; Nitschke, 1976). The parasympathetic nucleus of the intermediate nerve (nucleus parasympathticus n. intermedii) is located in the medulla dorsomedial to the somatic motor nucleus. These preganglionic axons leave the brainstem with the pars intermedia of the facial nerve. They join the main trunk of the facial nerve (Van Buskirk, 1945) and as they traverse the facial canal of the petrosal part of the temporal bone they leave as part of the chorda tympani nerve (Foley, 1945). The latter traverses the tympanic cavity and joins the lingual branch of the mandibular nerve. Synapse occurs in the mandibular ganglion (ganglion mandibulare) and the sublingual ganglion (ganglion sublinguale) and the postganglionic axons innervate the sublingual and mandibular salivary glands. The parasympathetic nucleus of the intermediate nerve also supplies glands of the tongue (see Chapter 7, The Tongue). See Figs. 15-1 and 15-2. The cell bodies of the preganglionic neurons that initially travel with the glossopharyngeal nerve are located in the medulla in the parasympathetic nucleus of the glossopharyngeal nerve (nucleus parasympatheticus n. glossopharyngei). This nucleus is located at the rostral end of the parasympathetic nucleus of the vagus nerve lateral to the hypoglossal nucleus and adjacent to the floor of the fourth ventricle. The preganglionic axons leave the medulla in the rootlets of the glossopharyngeal nerve, which enters the jugular foramen. These preganglionic axons branch from the glossopharyngeal nerve and become part of the tympanic plexus (plexus tympanicus) on the ventral surface of the petrosal portion of the temporal bone within the tympanic cavity. They leave the plexus in the minor petrosal nerve (n. petrosal minor) to synapse in the otic ganglion (ganglion oticum), which is adjacent to the external opening of the oval foramen and the origin of the mandibular nerve from the trigeminal nerve. The postganglionic axons run with the auriculotemporal nerve, a branch of the mandibular nerve to their destination on the gland cells of the parotid salivary gland. A large number of these postganglionic axons also run in the adventitia of the maxillary artery and a small number of axons run in the facial nerve (Holmberg, 1971). The zygomatic salivary gland also receives parasympathetic postganglionic axons from the otic ganglion. The otic ganglion is a plexiform structure containing a number of small ganglia and having close contact with the maxillary artery (Gienc & Kuder, 1983). (Figs. 15-1, 15-3, 15-4, and 15-6.) 2. Communicating rami from cervicothoracic ganglion to ventral branches of cervical and thoracic nerves 3. Left cervicothoracic ganglion 7. Left recurrent laryngeal nerve 8. Left tracheobronchial lymph node 13. Dorsal branch of vagus nerve 15. Ventral trunk of vagus nerve 16. Accessory lobe of lung (through caudal mediastinum) 17. Phrenic nerve to diaphragm 18. Paraconal interventricular a., v., and groove 20. Internal thoracic artery and vein The vagus nerve contains a number of functional components that include a large component of GVA axons and a smaller component of preganglionic parasympathetic GVE axons. The cervical portion of the vagus nerve is said to contain more than 80% “C” axons, with the balance myelinated (Fussey et al., 1973). The majority of the “C” axons are assumed to be afferent and the rest efferent parasympathetic fibers originating in the parasympathetic nucleus of the vagus nerve (nucleus parasympatheticus vagi). The paired nucleus is located in the dorsal part of the caudal medulla oblongata lateral to the hypoglossal nucleus ventral to the floor of the fourth ventricle. Various branches of the vagus are discussed in this section, but distribution of the branches other than preganglionic parasympathetic GVE axons is described in the section on cranial nerves. The vagal rootlets leave the brain along the dorsolateral sulcus of the medulla oblongata in series with the rootlets of cranial nerves IX and XI. Close association of some of the vagal axons with those of the accessory (XI) nerve for a very short distance has led to the description of these vagal axons as part of the cranial root of the accessory nerve in some species. At the level of the middle cervical ganglion (Fig. 15-5) the right recurrent laryngeal nerve (n. laryngeus recurrens) leaves the right vagus and passes dorsally around the caudal side of the right subclavian artery. As it runs cranially on the trachea, it lies deep to the common carotid artery in the angle between the longus colli muscle and the trachea. Near its origin, the right recurrent laryngeal nerve gives off a fairly large branch, the cervical cardiac nerve (n. cardiacus cervicalis), which receives contributions from the middle cervical ganglion, the vagal trunk, and the left recurrent laryngeal nerve. It is distributed mainly to plexuses along the right and left coronary arteries. The cervical cardiac nerve contains postganglionic sympathetic, preganglionic parasympathetic, and visceral afferent axons for the heart. Terminal parasympathetic ganglia in the heart contain neuronal cell bodies that differ markedly in size (Nonidez, 1939). As a consequence, the postganglionic axons also vary in diameter. These axons lie in the walls of the atria and the interatrial septum and form extensive cardiac plexuses. Smaller branches leave the plexuses and end as ring-shaped, club-shaped, or reticulated enlargements, which contact the surface of the cardiac muscle fibers. Similar enlargements may also occur along the course of the finer branchings. Terminations are especially abundant among the specialized muscle fibers of the sinoatrial (SA) and atrioventricular (AV) node. These nodes are also richly supplied with adrenergic fibers (Dahlström et al., 1965). Postganglionic parasympathetic axons of the cardiac ganglia are also distributed via nerve plexuses along branches of the coronary arteries. These axons form plexuses in the adventitia and finally end in relation to the smooth muscle cells of the outer media. Primarily, these axons innervate the arteries of the atrium and proximal part of the ventricles. Most of the parasympathetic distribution is to structures dorsal to the coronary sulcus, whereas sympathetic axons innervate the ventricles. Further information on intrinsic innervation of the heart may be found in Tcheng (1950, 1951), Holmes (1956, 1957), Uchizono (1964), Hirsch et al. (1964, 1965), Napolitano et al. (1965), Ehinger et al. (1967), Kent et al. (1974), Kyösola et al. (1976), Denn and Stone (1976), Geis et al. (1973), Martin et al. (1970), Coleridge et al. (1973), and Hopkins and Armour (1984). The main trunk of the right vagus continues caudally dorsal to the root of the lung. At this level it gives off several prominent branches along the bronchi (Ziemianski et al., 1967). Parasympathetic ganglia are found in the lung, and postganglionic parasympathetic axons have been traced to smooth muscle and glandular structures. The lung is also richly innervated with a wide variety of receptor structures as far distally as the alveoli. Receptors have been described in smooth muscle, tracheal and bronchial epithelium, respiratory bronchioles, alveolar ducts, and alveolar walls (Elftmann, 1943). The vagus supplies branches directly to the trachea and esophagus and also via the recurrent laryngeal nerve. Both dorsal and ventral vagal trunks supply branches to the esophagus before passing through the diaphragm at the esophageal hiatus. On reaching the abdominal cavity, the ventral vagus becomes plexiform for a short distance. Branches from this plexus supply mainly the liver and stomach. The hepatic branches (usually two or three) originate from both vagal trunks and run in the lesser omentum to the liver (Chiu, 1943). These pass between the caudate and left lateral lobe of the liver to form a simple plexus just rostral to the porta hepatis. Autonomic nerve branches are distributed from the plexuses to the cystic duct, gallbladder, left lateral lobe of the liver, and bile duct. Groups of axons also pass along the right gastric and cranial pancreaticoduodenal arteries to the pancreas and along the right gastric artery to the pylorus. A small filament may go directly to the duodenum. The second group of three or four gastric branches from the ventral vagus supply the parietal surface of the stomach. Some parasympathetic axons join the sympathetic plexus on the branches of the left gastric artery (Mizeres, 1955a). The dorsal vagal trunk supplies the cardiac region of the stomach and then forms a plexus on its visceral surface. Distribution is mainly to the lesser curvature and pyloric regions of the stomach. Branches of the dorsal vagal trunk may also join the celiac, left gastric, hepatic, and cranial mesenteric nerve plexuses associated with the corresponding arteries. Further references on abdominal distribution of the vagus nerve are those of Obrebowski (1965), Kapeller (1965), Mizeres (1955a), and Kemp (1973). Axons that make up the left recurrent laryngeal nerve leave the vagus at the level of the aortic arch and loop around the arch caudal to the ligamentum arteriosum. The recurrent laryngeal nerve then proceeds cranially along the left ventrolateral surface of the trachea adjacent to the ventromedial side of the esophagus. Branches are supplied to the trachea and esophagus as the nerve passes to its termination in the larynx (Hwang et al., 1948; Watson, 1974). In the cervical region the left recurrent laryngeal supplies more branches to the esophagus than it does to the trachea. The opposite is true of the right recurrent laryngeal nerve. In dogs whose esophagus lies to the right of the midline in the cervical region, there is a more even distribution of branches of right and left recurrent laryngeal nerves to both trachea and esophagus. Parasympathetic ganglionic neurons may be found scattered in the walls of structures that they supply or else as part of well-defined plexuses in similar regions. These intramural nerve plexuses are most prominent in the digestive tract (Filogamo, 1950; Lawrentjew, 1931; Richardson, 1960; Schofield, 1961). The myenteric plexus (plexus myentericus), is located between the longitudinal and circular muscle layers, and the submucosal plexus (plexus submucosus) lies in the submucosa of the digestive tube (El’bert, 1956; Leaming & Cauna, 1961). Ganglia are common in these enteric plexuses (plexus entericus). Prominent ganglionated plexuses are also found in the atria of the heart. Much of their regulating effect is thought to be mediated by way of the SA and AV nodes.
The Autonomic Nervous System
Parasympathetic Division
Oculomotor (III)
Facial (VII)
Glossopharyngeal (IX)
Vagus (X)
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


