Chapter 11 Reproductive system
Male reproductive system
The male dog is known as a dog; the male cat is known as a tomcat. The reproductive system of the dog and the tomcat are generally similar – any differences will be described where appropriate (Figs 11.1, 11.2). The parts of the male reproductive tract are:
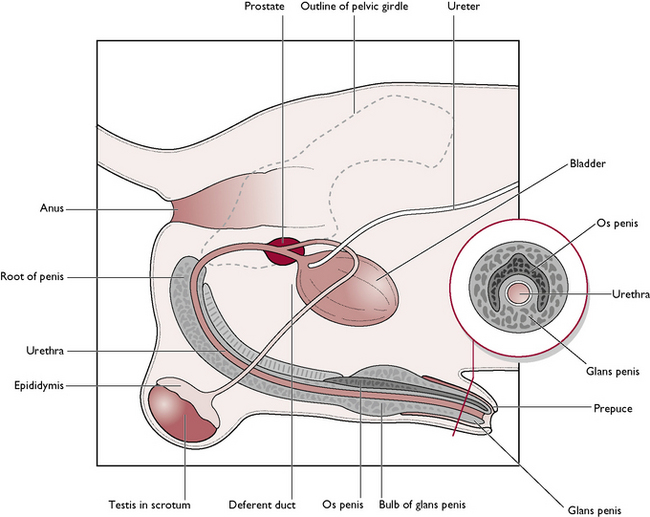
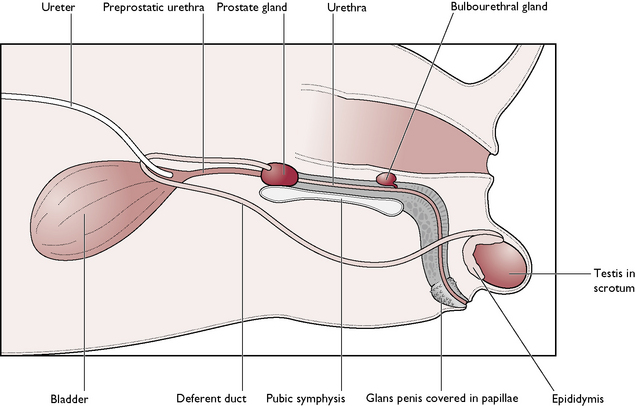
The testis
The testis is the male gonad, with functions as follows:
Each testis is an oval-shaped structure wrapped in a double layer of peritoneum known as the tunica vaginalis (Fig. 11.3). The testicular tissue consists of numerous blind-ending tubules known as seminiferous tubules which are lined by two types of cells:
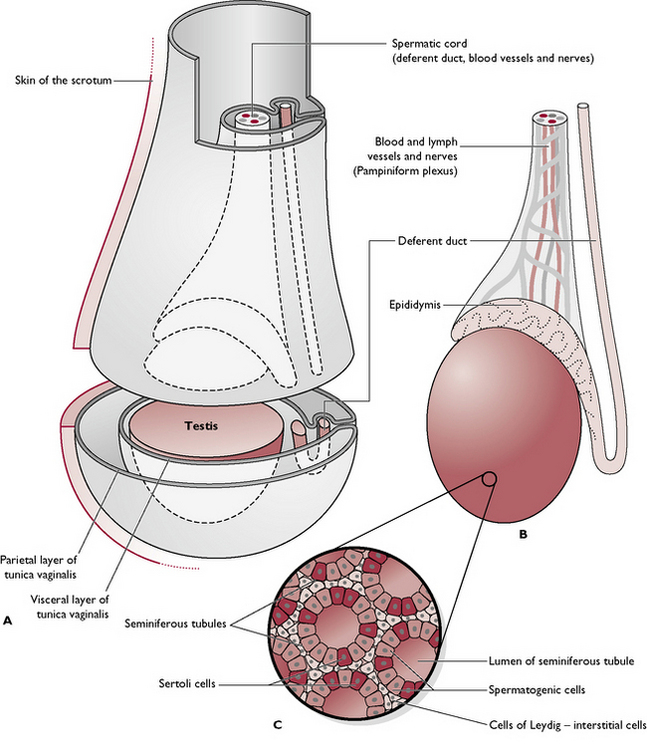
Fig. 11.3 A Testis within scrotum. B Scrotum removed. C Cross-section through the seminiferous tubules.
Lying between the tubules are the cells of Leydig or interstitial cells. They secrete testosterone and are under the control of interstitial cell stimulating hormone (ICSH) (see Ch. 6), produced by the anterior pituitary gland.
The coiled seminiferous tubules make up most of the testicular tissue and eventually combine to form slightly larger efferent ducts. These drain into the epididymis, lying along the dorsolateral border of the testis. The cauda epididymis or tail is attached to the caudal extremity of the testis and is the point at which the temperature of the testis is lowest. It is here that sperm are stored and undergo a period of maturation ready for fertilisation (Fig. 11.4).
Testicular descent
In the early embryo, the undifferentiated gonads develop inside the abdomen close to the kidney. In the male, the gonad becomes the testis and a band of tissue known as the gubernaculum forms and runs from the caudal end of each testis to the inside of the developing scrotal sac. During late gestation, the testes are pulled caudally by the contraction of the gubernaculum and they migrate through the abdomen. The testes leave the abdominal cavity viathe inguinal canal – a channel between the fibres of the external abdominal oblique muscle in the groin or inguinal area (see Ch. 4). As each testis with its associated blood capillaries, nerve and deferent duct passes through the inguinal canal into the scrotum, it becomes wrapped in a double fold of peritoneum which forms the tunica vaginalis (Fig. 11.3).
Deferent duct
The epididymis continues as the deferent duct (also called the vas deferens or ductus deferens; Figs 11.1, 11.2), which passes out of the scrotum into the abdominal cavity via the inguinal canal within the spermatic cord. The spermatic cord is wrapped in the tunica vaginalis and also contains the testicular artery and vein and the testicular nerve. Lying within the cord is a strip of muscle derived from the internal abdominal oblique muscle and known as the cremaster muscle. Contraction of this muscle raises the testis closer to the body in response to cold and works in conjunction with the Dartos muscle to maintain a constant temperature for the testes.
During ejaculation, the sperm and fluid produced in the seminiferous tubules are propelled along the epididymis and up the deferent duct, which joins the urethra. At this junction, the walls of the deferent ducts are thickened and glandular; the whole area is surrounded by the prostate gland (Figs 11.1, 11.2).
The penis
The functions of the penis are to:
The urethra runs through the centre of the penis and extends from the bladder to the tip of the penis. It is shared by both the reproductive and urinary systems (see Ch. 10). The penis of the dog and the cat are anatomically different.
The dog
The penis runs from the ischial arch of the pelvis, passes cranioventrally along the perineum and between the hind limbs (Fig. 11.5). The urethra lies in the centre and is surrounded by a layer of cavernous erectile tissue known as the corpus spongiosum penis. This is expanded proximally into the bulb of the penis and, towards the tip, as the glans penis. Surrounding this and serving to attach the penis to the ischial arch is a pair of erectile tissue crura (sing. crus), known as the corpus cavernosum penis. These form the root of the penis at its attachment to the ischial arch. The urethra runs in a groove between the two crura.
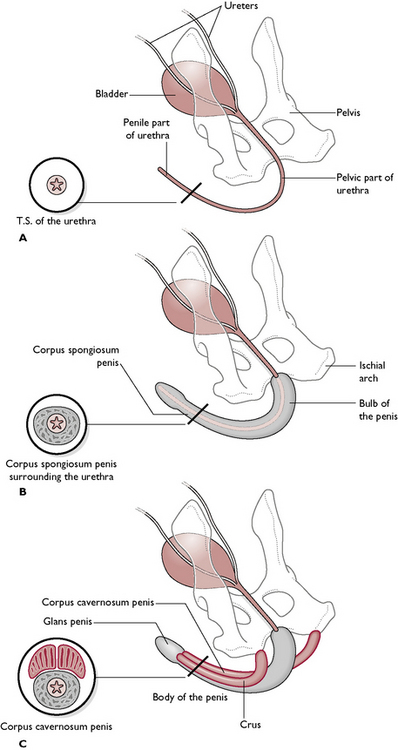
The distal part of the penis is contained within a sheath of hairy skin known as the prepuce. This is suspended from the ventral abdominal wall and covers and protects the penis. It is lined with mucous membrane and is well supplied with lubricating glands. During mating, the prepuce is pushed back to reveal the glans penis. Afterwards, the retractor penis muscle pulls the penis back into the prepuce.
The tomcat
The main parts of the penis are similar to those of the dog, except that the cat penis is shorter and points backwards – the external opening is ventral to the anus (Fig. 11.2). The glans penis is covered with tiny barbs, which elicit a pain reflex as the male withdraws from the female after mating. This stimulates the nerve pathway to the hypothalamus, resulting in ovulation approximately 36 hours later – known as induced ovulation. The os penis lies ventral to the urethra in the cat. During sexual excitement, the penis engorges and points cranioventrally so that the mating position in cats is similar to that seen in the dog.
Accessory glands
The function of the accessory glands is to secrete seminal fluids which:
There are two types of gland (Figs 11.1, 11.2):
Female reproductive system
The reproductive tract of the bitch and queen are similar and vary only in size (Fig. 11.6). The tract is designed to carry several fetuses during a single pregnancy and is said to be bicornuate (two horns). The bitch and the queen bear litters of young: they are multiparous.
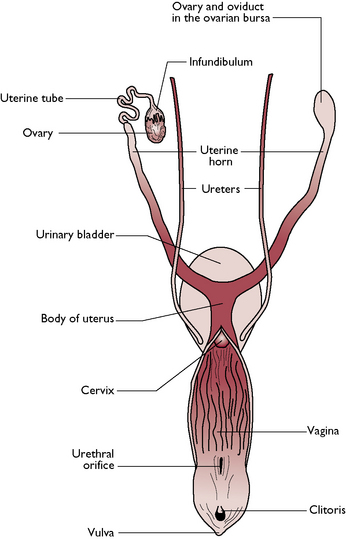
Fig. 11.6 Dorsal view of the reproductive system of the bitch.
(With permission from Colville T, Bassett JM 2001 Clinical anatomy and physiology for veterinary technicians. Mosby, St Louis, MO, p 330.)
The ovary
The ovary is the female gonad. The functions of the ovary are:
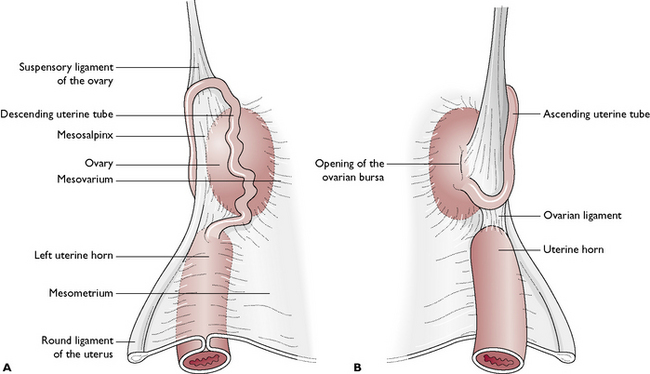
There is a pair of ovaries, one lying on each side of the dorsal abdominal cavity, caudal to the kidney (see Ch. 10, Fig. 10.2). The ovary is held close to the kidney by the ovarian (suspensory) ligament (Fig. 11.7). The ovary is suspended from the dorsal body wall by part of the visceral peritoneum called the mesovarium, which also encloses the infundibulum of the uterine tube. Part of the mesovarium forms a pocket-like structure known as the ovarian bursa, which completely covers the ovary. Within this is a small opening allowing ova to leave the ovary – this is a potential means of entry of infection into the peritoneal cavity.
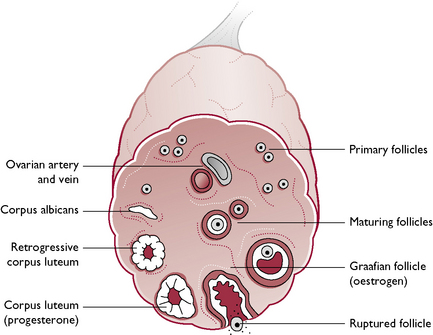
The tissue of the ovary consists of a framework of connective tissue, smooth muscle and blood capillaries, within which are a large number of germ cells and developing follicles (Fig. 11.8). In an immature animal, each ovary is oval with a smooth outline but, as sexual maturity approaches, the ovary becomes nodular as the follicles enlarge.
The uterine tube
This is also known as the oviduct or Fallopian tube (Fig. 11.6). The functions of the uterine tubes are:
Each uterine tube is a narrow convoluted structure lying close to the ovary (Fig. 11.7). The open end is funnel-shaped and known as the infundibulum. It is fringed with finger-like processes known as fimbriae, which spread over the surface of the ovary to capture ova as they are released. The ova pass down the lumen of the tube, which is lined with ciliated columnar epithelium. The cilia propel the ova along the tube towards the uterine horns. The uterine tube is suspended by part of the visceral peritoneum known as the mesosalpinx.
The uterus
The uterus is a Y-shaped structure lying in the midline of the dorsal abdomen (Fig. 11.6). During pregnancy, the weight of the conceptuses pulls the uterus ventrally and at full term it occupies the greater part of the abdomen. The function of the uterus is:
The wall of the uterus has three layers:
The cervix
The cervix is a short, thick-walled muscular sphincter that connects the uterine body with the vagina (Fig. 11.6). Running through the centre is a narrow cervical canal, which is normally tightly closed and relaxes only to allow the passage of sperm or fetuses. During pregnancy, the canal is blocked by a mucoid plug, which protects the conceptuses from infection. In the non-pregnant animal, the cervix lies in the pelvic cavity but during pregnancy the weight of the conceptuses pulls the cervix cranially and ventrally over the edge of the pelvic brim.
The vulva
The vulva marks the external opening of the urogenital tract. It consists of two parts:
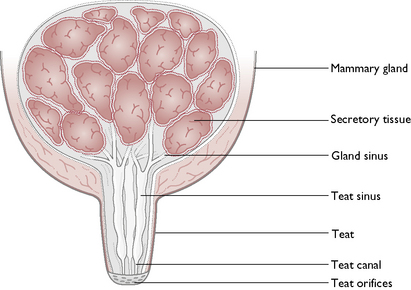
The mammary glands
Each gland consists of glandular tissue embedded in connective tissue and lined by a secretory epithelium (Fig. 11.9). The milk produced drains through a network of sinuses that eventually form teat canals. These open on to the surface of each teat, known as a teat orifice. Each gland has one teat but each teat has several orifices.
Lactation
Composition of milk
This varies between different species and is an important consideration when feeding orphaned animals. The milk produced by the bitch and the queen is more concentrated and contains more protein and twice as much fat as cow’s milk. The average composition of milk is shown in Table 11.1.
Table 11.1 Average composition of milk
| Constituent | Quantity (%) |
|---|---|
| Water | 70–90 |
| Fat | 0–30 |
| Protein | 1–15 |
| Carbohydrate | 3–7 |
| Minerals | 1.5–1: calcium phosphate, magnesium, sodium, potassium and chlorideMilk is deficient in iron and copper; traces of iodine, cobalt, tin and silica are present |
| Vitamins | A, B2, B5, E, KMilk is low in vitamins C and D |
NB. Milk from a cat contains the amino acid taurine, as cats have a specific requirement for this.
The oestrous cycle
The oestrous cycle is the rhythmic cycle of events that occurs in sexually mature non-pregnant female mammals and includes limited periods of sexual receptivity known as oestrus. The function of the oestrous cycle is:
< div class='tao-gold-member'>
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


