Chapter 24. Performance
ENDURANCE PERFORMANCE
Endurance performance in dogs, as in humans, involves prolonged periods of exercise at sub-maximal levels of exertion. Much of what we currently understand about the nutritional needs of hard-working endurance dogs comes from studies of Alaskan sled dogs. Sled dogs working in cold environments represent the ultimate canine endurance athletes. These dogs have been used as the accepted mode of transportation in Alaska and surrounding territories for many years. During the Gold Rush of the late 1800s, prospectors traveled to the Yukon Territory and used dogs to cross the northern tundra in search of gold. As mining camps and towns developed, sled dog trails were forged to connect coastal towns to interior camps. These trails provided the major transport routes for the provision of food and supplies to the camps. Although most of the mining camps are now mere ghost towns, dogs and dog sledding remain an integral part of the area’s economy and culture. In addition to their importance in Alaska, sled dog teams have been used by explorers of the North and South Poles. Robert Perry reached the North Pole via dog sled in 1909, and Roald Amundsen reached the South Pole with his team of dogs in 1911. Sled dog teams continue to provide transportation in certain areas of the world. In addition, mushing and sled dog racing have become popular outdoor sports for many dog enthusiasts. Today, sled dog races are held in many parts of the world, including Europe, Alaska, and several of the lower 48 states.
The Iditarod is one of the most well-known and publicized long-distance sled dog races. Informally called the “the last great race on earth,” the Iditarod is run every March, beginning in Anchorage and ending in Nome. The trail that is followed is symbolically designated to be 1049 miles long (1000 miles because it is at least that long, and 49 miles because Alaska was the forty-ninth state to be admitted to the union). Officially, the race distance is 1158 miles by the northern route or 1163 miles by the southern route. Because of its great length and the difficult terrain, the Iditarod represents the ultimate endurance test for the canine athlete. Dog teams average up to 100 miles per day in subzero temperatures and over difficult snow-covered terrain. Nutritionists and exercise physiologists interested in the care and nutrition of these dogs have used this race and similar long-distance sled dog races to collect valuable information about the energy, nutrient, water, and electrolyte needs of working dogs. In addition, studies of certain types of hunting dogs, search and rescue dogs, and military dogs also provide information regarding the needs of dogs that work for long periods of time at submaximal levels of exertion. This research has provided information for the development of diets that provide optimal nutrition for working dogs; it has also contributed to improved information about the proper care and husbandry of dogs engaged in endurance exercise.
Physiology
The type of work that occurs during endurance competitions differs from that performed during short races or sprinting events, such as Greyhound racing or lure coursing. Greyhounds engage in brief, intense bouts of high-speed running, while sled dogs pull for several hours at a time at slower speeds. Dogs used for hunting typically engage in both types of exertion. These dogs often work for hours at a time over a period of several days (endurance), interspersed with occasional bouts of high-intensity sprinting. Metabolically, the energy necessary for short and intense sprints is obtained primarily through anaerobic pathways and secondarily through aerobic metabolism. In contrast, the energy that is needed for endurance work is predominantly derived from aerobic metabolism.
Greyhounds engage in brief, intense bouts of high-speed running, sled dogs pull for several hours at a time at slower speeds, and dogs used for hunting typically engage in both types of exertion. Metabolically, the energy necessary for short and intense sprints is obtained primarily through anaerobic pathways and secondarily through aerobic metabolism. In contrast, the energy that is needed for endurance work is predominantly derived from aerobic metabolism.
Fat and carbohydrate are the two principal fuels that supply energy to working muscle. During the low-intensity (aerobic) exercise associated with endurance, fat is the most important fuel used. This is supplied by free fatty acids (FFAs) derived from triglycerides of muscle and other tissues. As the intensity of exercise increases, a shift toward more anaerobic metabolism occurs, and carbohydrate becomes increasingly important as a source of energy. This carbohydrate is supplied by muscle glycogen and, as muscle glycogen becomes depleted, is supplemented by the production of glucose by the liver (gluconeogenesis). This metabolic response to different intensities of exercise and training has been observed in rats, humans, and dogs. 1.2.3.4. and 5. For example, the rate of muscle glycogen utilization in trained dogs completing a high-intensity anaerobic test is almost twenty-fold greater than the rate observed in dogs during a period of aerobic exercise. 6
Fat is the most important fuel used during low-intensity, aerobic exercise. This is supplied by free fatty acids derived from triglycerides of muscle and other tissues. As the intensity of exercise increases, a shift toward more anaerobic metabolism occurs, and carbohydrate becomes increasingly important as a source of energy. This carbohydrate is supplied by muscle glycogen and via gluconeogenesis.
An examination of the types of muscles that are involved in endurance exercise provides information about the types of energy sources needed. The skeletal muscles of dogs contain three main types of muscle fibers: type I (slow twitch) and types IIa and IIb (fast twitch). Slow-twitch fibers have a high capacity for aerobic metabolism, and fast-twitch fibers can use both oxidative (aerobic) and anaerobic pathways. 7 The slow-twitch fibers use fatty acids and glucose for fuel and are believed to be important for endurance events. Although these fibers are found in all skeletal muscles, their numbers predominate in antigravity muscles such as the dog’s anconeus and the quadratus muscles. 8 In general, endurance athletes have higher numbers of well-developed, slow-twitch fibers, and athletes involved in high-speed sprinting events have a higher proportion of fast-twitch fibers. Athletic conditioning is important for conditioning muscles and building muscle mass. Athletic training for endurance events increases the body’s reliance upon fatty acid oxidation during submaximal bouts of exercise. This adaptation has been demonstrated in rats, humans, and dogs. 1. and 2. Moreover, the type of diet that is fed affects the use of fatty acids by working muscles, independent of the effects of training. 6. and 9.
Glycogen Loading
Although it is generally accepted that energy is the nutrient of most concern for working dogs, there has been much debate about the best way to supply dietary energy to enhance endurance performance. Increasing stamina and strength are goals for the nutritional programs of many human athletes. As a result, a great deal of research has been conducted concerning ways to supply fuel to long-distance runners and cyclists. 1. and 10. An important limiting factor in prolonged exercise is the amount of glycogen present in the working muscles because the onset of fatigue is highly correlated with muscle glycogen depletion. 11. and 12. In addition, endurance for submaximal exercise can be increased by raising muscle glycogen stores and decreased by lowering muscle glycogen. 12. and 13. Therefore a major goal when feeding human endurance athletes is to either increase muscle glycogen stores or delay muscle glycogen depletion during periods of exercise.
The procedure of glycogen loading (also called carbohydrate loading) was developed for human athletes with the intent of increasing muscle glycogen stores before periods of prolonged exercise. Glycogen loading was initially accomplished by first depleting muscle glycogen through exhaustive exercise and/or consumption of a low-carbohydrate diet, followed by the consumption of a high-carbohydrate diet for 4 to 7 days. 14. and 15. The preliminary glycogen depletion phase presumably resulted in glycogen supercompensation when the subject consumed a diet that is high in starch for several subsequent days. The beneficial effects that higher initial glycogen stores have on endurance are believed to be the result of the availability of larger amounts of glycogen for anaerobic energy metabolism in the working muscles. However, controlled studies have shown that in the first stage, glycogen depletion is unnecessary and may even be detrimental to training performance and health. 16. and 12. Today, many endurance athletes regularly consume a high-carbohydrate diet and concentrate on carbohydrate loading for the last few days prior to an endurance event but do not engage in the preliminary depletion phase.
Providing Energy for Working Dogs: Fat or Carbohydrate?
There are important differences between human athletes and working dogs. In dogs, approximately 70% to 90% of the energy for sustained work is derived from fat metabolism, and only a small amount of energy is derived from carbohydrate metabolism. 17. and 18. Similarly, early field studies with sled dogs and laboratory studies with Beagles indicated that the ability to use fatty acids through aerobic pathways was more important than the use of muscle glycogen through anaerobic pathways during hard work. 1.19. and 20. In an often referenced study, a dog team that was fed a high-carbohydrate diet performed poorly and developed a stiff gait while racing. 21 When the dogs were changed to a diet containing increased levels of fat and protein, performance improved and the observed lameness resolved. The researchers suggested that feeding a high-carbohydrate diet to sled dogs may be responsible for the occurrence of a form of exertional rhabdomyolysis. Exertional rhabdomyolysis is a disorder caused by rapid anaerobic metabolism of muscle glycogen, resulting in an accumulation of lactic acid. Lactic acid accumulation can have several adverse side effects, including damage to muscle-tissue membranes, edema, and inhibition of lipolysis and glycolysis. These studies were the impetus for the theory that dietary fat is metabolically the preferred fuel for hard-working dogs.
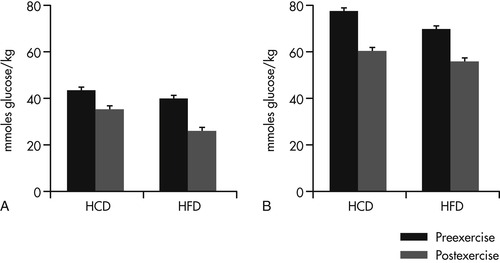
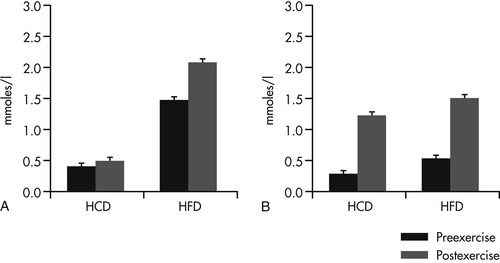
Subsequent controlled studies with several types of working dogs have supported this hypothesis and have also provided important information regarding the feeding management and training for working dogs. Both the level of conditioning and the type of diet fed significantly affect energy use in working dogs. In a study with 16 Alaskan Huskies, one group of dogs was fed a high-fat diet containing 60% of calories as fat, while the second group was fed a high-carbohydrate diet, containing 60% of calories as carbohydrate. 22 The dogs completed standard aerobic and anaerobic tests on a treadmill prior to conditioning, after a 4-week aerobic training period, and again after a 4-week anaerobic training period. Results of the aerobic tests showed that both groups of dogs relied primarily upon FFAs and used little glycogen to complete the tests (Figure 24-1 and Figure 24-2). Dogs that were fed the high-fat diet had significantly greater preexercise and postexercise FFA concentrations when compared with dogs fed the high-carbohydrate diet, even before they were conditioned through training. The concentration of FFAs in the blood is a major determinant of fatty acid utilization by muscle, and it is an accepted indicator of fatty acid metabolism. These results showed that feeding a high-fat diet to working dogs before and during athletic training enhances the dog’s ability to mobilize and use fatty acids as fuel.
 |
| Figure 24-1 (From Reynolds AJ, Taylor CR, Hoeppler H, and others: The effect of diet on sled dog performance, oxidative capacity, skeletal muscle microstructure, and muscle glycogen metabolism. In Carey DP, Norton SA, Bolser SM, editors: Recent advances in canine and feline nutritional research, Iams international nutritional symposium, Wilmington, Ohio, 1996, Orange Frazer Press.) |
 |
| Figure 24-2 (From Reynolds AJ, Taylor CR, Hoeppler H, and others: The effect of diet on sled dog performance, oxidative capacity, skeletal muscle microstructure, and muscle glycogen metabolism. In Carey DP, Norton SA, Bolser SM, editors: Recent advances in canine and feline nutritional research, Iams international nutritional symposium, Wilmington, Ohio, 1996, Orange Frazer Press.) |
A second controlled study examined the effect of diet on oxygen consumption (V o2 max), mitochondrial volume, and maximal rates of fat oxidation in a group of endurance-trained Labrador Retrievers. 9 When dietary fat was increased from 15% to 60% of calories, dogs showed almost 50% increase in V o2 max and a 45% increase in maximal fat oxidation during aerobic exercise tests. These values indicate enhanced efficiency of fat utilization and capacity for aerobic work. Mitochondrial volume in muscles increased 50% in response to increasing fat in the diet. There is speculation that feeding a high-fat diet improves aerobic work capacity and efficiency in dogs by stimulating mitochondrial growth.
Although fat is the primary metabolic fuel for endurance dogs, and high-fat diets best supply this fuel, adequate muscle glycogen stores are still important during exercise. As in other species, muscle glycogen depletion during prolonged exercise is associated with fatigue and a decline in performance in working dogs. The two factors that appear to have the greatest influence on muscle glycogen stores are athletic conditioning and diet (see Figure 24-1). In the study described previously, only small amounts of glycogen were used by dogs during the aerobic tests. 9 However, some glycogen metabolism is always necessary to permit the continuation of FFA metabolism during aerobic work. In addition to this need, racing sled dogs experience periods of intense running near the end of races or when covering difficult terrain. Similarly, hunting dogs and certain types of service and sports dogs work predominantly at low intensities, interspersed with occasional bouts of high-intensity physical exertion.
To learn more about the needs of dogs during these periods, researchers also examined energy metabolism in endurance-trained sled dogs during bouts of intense anaerobic activity. 6 Completion of an initial 4-week training period significantly increased the muscle glycogen levels in all of the dogs in the study, regardless of the type of diet that was fed. While dogs that were fed a high-carbohydrate diet stored more muscle glycogen than those fed a high-fat diet, the carbohydrate-fed dogs also metabolized more glycogen to complete the anaerobic test. As a result, muscle glycogen levels at the completion of the standard anaerobic test were depleted similarly in the groups of dogs fed the two different diets. As in other species, dogs performing anaerobic tests rely heavily upon muscle glycogen as an energy source. The researchers concluded that feeding high-fat diets to endurance-trained dogs not only prepares muscles to efficiently mobilize and use FFAs as an energy source, it also appears to have a glycogen-sparing effect that helps to prolong glycogen use.
Finally, although dogs react similarly to other species by showing rapid glycogen depletion following a single bout of strenuous exercise, there is recent evidence that trained sled dogs are resistant to prolonged glycogen depletion when subjected to repeated exercise episodes over multiple days. Early studies with human athletes suggested that completing consecutive days of prolonged running results in cumulative muscle glycogen depletion, and it is this depletion and the inability to completely replenish glycogen stores that is responsible for the onset of fatigue and loss of performance. 23. and 24. Continuing to exercise when muscle glycogen stores are low has also been associated with increased protein degradation in skeletal muscle, a potential risk factor for injury. 25 Recent studies with a group of trained Alaskan sled dogs examined muscle glycogen levels during prolonged bouts of exercise. In the first study, 36 conditioned sled dogs consuming a high-fat, low-carbohydrate diet ran approximately 100 miles per day for up to 5 consecutive days. 26 Muscle glycogen was measured prior to the start of the exercise period and immediately following each day’s run. The first day’s workout caused a rapid depletion in glycogen levels to about 21% of preexercise levels. However, glycogen levels subsequently increased to 50% to 65% of initial stores after the second day of exercise and remained at that level for the remainder of the 5-day period. This change was especially remarkable since the dogs were consuming relatively little carbohydrate in their training diet. The underlying metabolic cause for muscle glycogen replenishment may have been enhanced use of noncarbohydrate fuel sources during subsequent days of exercising or to the rapid replenishment of glycogen stores during rest periods. Although the study was not designed to determine the underlying cause, serum ketones and glycerol concentrations increased over the 5-day period, suggesting that utilization of fat-based energy sources was at least partially responsible for the maintenance of muscle glycogen in the exercising dogs. 27
A subsequent study was conducted to determine the underlying cause of glycogen replenishment in dogs during multiple days of endurance work. 28 Changes in muscle glycogen and muscle triglyceride concentrations were measured in a group of sled dogs over a 4-day period of 100-mile per day training runs. A group of six similarly trained dogs served as the nonexercising control group. All of the dogs were fed a high-fat training diet and muscle biopsies were performed immediately after each day’s run and again following feeding and a period of rest. Following the first day’s run, dogs had utilized more than 60% of the glycogen stored in skeletal muscle, corroborating the results of the previous study. However, running the same distance on each subsequent day resulted in only a 5% additional decrease in glycogen stores. This change diminished to almost negligible levels on the final day. In contrast, as the use of muscle glycogen decreased, the use of muscle triglyceride increased. These findings are supported by previous work showing that extramuscular fat and protein substrate play an important role in supporting work in sled dogs during periods of prolonged and submaximal exercise. It appears that plasma FFAs become the predominant energy substrate during this time. Because blood urea nitrogen (BUN) concentrations also increase during prolonged and submaximal exercise, protein substrates may also be important. Together the results of these studies suggest that the primary metabolic adaptation that occurs during prolonged and repeated bouts of exercise in dogs is a reduced rate of glycogen depletion caused most likely by increased reliance on fat-related energy substrates, most importantly plasma FFAs. Although there is some evidence that human athletes subjected to multiple days of endurance exercise also shift their metabolism away from muscle glycogen and toward fat-based substrates, the dog may be especially efficient at this type of shift given the dog’s naturally high aerobic metabolic capacity and demonstrated response to training and to high-fat diets. 29.30. and 31.
Both the level of conditioning and the type of diet fed affect energy use in working dogs. Feeding a high-fat diet to working dogs before and during athletic training enhances the dog’s ability to mobilize and use fatty acids as fuel during endurance exercise and may help to spare glycogen stores from depletion.
Protein for Working Dogs
Endurance training and racing result in increased protein needs for dogs. 32 Several physiological and metabolic changes contribute to an increased dietary requirement. Athletic conditioning results in adaptive anabolic changes that facilitate efficient delivery of oxygen and nutrients to working muscles. These changes include increases in blood volume, red blood cell mass, capillary density, mitochondrial volume, and the activity and total mass of metabolic enzymes. 33 The increased tissue mass associated with athletic training must be supplied by additional protein in the diet. In addition, there is a slight but significant increase in protein catabolism during endurance exercise associated with an increased rate of skeletal muscle turnover. 34. and 35. This occurs as a result of exercise-induced muscle damage and oxidation of protein. It is estimated that up to 10% of energy in exercising dogs can be derived from the metabolism of gluconeogenic amino acids. 36. and 37.
In a study of the protein needs of dogs during endurance training, diets supplying either 16%, 24%, 32%, or 40% of calories from protein were fed to sled dogs throughout a 12-week training period. 38 Dogs that were fed 40% protein maintained a larger plasma volume and red blood cell mass during training than dogs fed diets containing less than 40% protein. The amount of protein in the diet also appeared to influence susceptibility to injury. While there were no injuries in any of the dogs that consumed the 32% or 40% protein diets, all of the dogs fed the 16% diet were injured at some point during the 12-week period, which kept them out of training for 2 or more days. It appears that the proportion of energy that is supplied by protein should be increased in the diets of endurance dogs in training to ensure adequate tissue accretion, prevent tissue loss, and possibly aid in the prevention of injury. Optimal protein concentrations of between 30% and 40% of calories are recommended in foods formulated for endurance dogs (Box 24-1).
BOX 24-1
Feed a highly digestible, energy- and nutrient-dense diet (4000 kcal of metabolizable energy/kg or greater).
Diet should contain:
Calories from protein: 30% to 35%; high-quality animal source proteins
Calories from fat: 50% to 65%
Calories from carbohydrate: 10% to 15%
Omega-6:omega-3 fatty acid ratio between 5:1 and 10:1
Moderately fermentable fiber: 3% to 7%
Provide continual access to clean, fresh water.
Feed two or more meals per day on a portion-controlled basis.
Feed the largest meal of the day after the day’s training is complete.
Provide a meal 1.5 to 2 hours before training or an endurance event.
Feed a carbohydrate-containing supplement immediately after endurance exercise to promote glucose repletion.
While providing enough protein in the diet is important, feeding a diet that contains more protein than the dog needs is neither necessary nor beneficial. Protein consumed in excess of the needs of tissue replacement and growth will be used as an energy source. Protein is one of the least desirable muscle energy fuels because it is inefficiently metabolized and cannot be stored in the body like fat or glycogen. The diet should supply adequate calories as fat and carbohydrate so that the protein that is fed can be used primarily for tissue protein synthesis and spared for use as an energy source. For the canine athlete, the most efficiently used fuel appears to be fat, with carbohydrate supplying a smaller proportion of the calories in the diet.
Performance dogs have increased needs for dietary protein. Athletic conditioning leads to increases in blood volume, red blood cell mass, capillary density, and mitochondrial volume. The increased tissue mass associated with this training must be supplied by additional protein in the diet. In addition, there is a slight but significant increase in protein catabolism during endurance exercise associated with an increased rate of skeletal muscle turnover. Finally, it is estimated that up to 10% of energy in exercising dogs can be derived from the metabolism of gluconeogenic amino acids.
Caloric Requirement during Endurance Work
The total energy requirement of an endurance dog depends on the intensity and duration of the exercise and the environmental conditions in which the animal is working. Early research suggested that energy needs increase to between 1.5 and 2.5 times the normal maintenance requirements in dogs working in ambient temperatures. 39.40. and 41. Working in cold weather may further increase requirements by about 50%. Data for these estimates were collected from draft dogs that were covering relatively short distances and traveling at slower speeds than racing sled dogs. Typical dog teams used for hauling freight travel less than 20 miles per day at speeds of about 5 miles per hour. Energy needs for these dogs are estimated to be between 4000 and 8000 kilocalories (kcal)/day. 41. and 42.
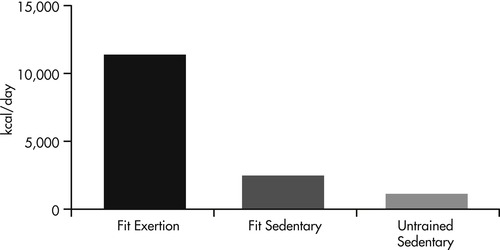
In contrast, dogs trained for racing in long-distance sled races typically run 70 miles or more per day and travel at speeds of up to 9 miles per hour. Studies of these dogs found that racing sled dogs competing in a medium distance race (300 miles) expended an average of 11,200 kcal/day and consumed an average of 10,600 kcal/day (Figure 24-3). 43 These values are equivalent to burning 460 kcal/kilogram (kg) of body weight per day and consuming 440 kcal/kg of body weight per day. The researchers observed that the calculated values for sustained metabolic rate for these dogs appears to exceed previously predicted maximal values for mammals of their size. These extraordinarily high energy requirements and expenditures predicate the need to provide a diet that is energy dense and highly digestible to allow the ingestion of needed calories in a volume of food that the dog’s stomach and gastrointestinal system are capable of handling. However, the vast majority of working dogs do not work for the duration and at intensities that result in such dramatic increases. Although a limited number of studies have been conducted, measured estimates for hunting dogs fall between the earlier estimates provided of 1.5 and 2.5 times maintenance requirements (see p. 255).
 |
| Figure 24-3 (From Hinchcliff KW, Reinhart GA, Burr JR, and others: Energy metabolism and water turnover in Alaskan sled dogs during running. In Carey DP, Norton SA, Bolser SM, editors: Recent advances in canine and feline nutritional research, Iams international nutritional symposium, Wilmington, Ohio, 1996, Orange Frazer Press.) |
Water and Electrolyte Requirements
The same researchers who measured caloric intake and expenditure in racing sled dogs also examined water intake, water turnover, and electrolyte balance during medium- and long-distance races. 44.45. and 46. Dogs competing in a 300-mile race had a measured water turnover rate of approximately 250 milliliters (ml)/kg of body weight per day. This is equal to about 5 liters (L) per day for an average-size sled dog. By comparison, sled dogs that were not racing had water turnovers of only 0.9 L per day. Dogs lose water primarily through respiration, urine, feces, and, to a very small degree, perspiration. The extremely high water loss in working sled dogs is primarily a result of increased urinary water loss. In all animals, urinary water loss is related to the obligatory urinary solute load, which in turn is affected by the quantity of food consumed and the composition of the diet. 47 Diets with a high protein concentration and/or large caloric intake result in higher obligatory solute loads. Because sled dogs consume up to 12,000 kcal/day when they are racing, they have extremely high solute loads and, as a result, very high obligatory urinary water losses. Dogs must be provided with fresh water frequently during racing to offset these losses and prevent dehydration.
< div class='tao-gold-member'>
Only gold members can continue reading. Log In or Register to continue
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


