, Monika Hassel2 and Maura Grealy3
(1)
Centre of Organismal Studies, University of Heidelberg, Heidelberg, Germany
(2)
Spezielle Zoologie, Universität Marburg FB Biologie, Marburg, Germany
(3)
Pharmacology and Therapeutics, National University of Ireland Galway, Galway, Ireland
20.1 “A Second Embryogenesis” Creates a Second Phenotype
20.1.1 Most Amphibians and Invertebrates Undergo Major Phenotypic Transformations Changing from a First to a Second Elaborated Form
Metamorphosis (Greek: transformation, change of form) refers to fundamental remodelling of the whole body, and is associated with a fundamental change in the mode of life. The new phenotype occupies a new ecological niche and colonizes a different habitat. Larvae and adults utilize different nutritional resources and often live in different environments.
Most benthic marine invertebrates produce planktonic larvae. These free-living stages enable the species to first make use of planktonic prey and subsequently to exploit a scattered or transient ecological niche suited for the adult phase. Planktonic invertebrate larvae include those of echinoderms, such as the pluteus of sea urchins (Figs. 4.1 and 4.2); the trochophore of the polychaete annelids (sandworms, tube worms; Fig. 4.22); the veliger larvae of molluscs; the actinotrocha of the phoronids (slender worm-like sedentary creatures); the nauplius of the crustaceans (lobsters, crabs, barnacles). Planktonic larvae enter the adult population through metamorphosis and settlement. The planula larvae of the Cnidaria are also free-living. They undergo metamorphosis into the sessile stage of polyps, as do the tadpole-like larvae of the tunicates which also undergo metamorphosis into sessile phenotypes.
In insects and amphibians, the acquisition of a new phenotype in metamorphosis implies the transition to a different habitat or at least a change in the nutritional resources that are exploited. Though the amphibians were the first vertebrates to spend a significant portion of their lives in terrestrial habitats, most present-day amphibians still return to the water to reproduce. The larvae live in water and feed preferentially on algae and plants. The terrestrial adults are carnivorous. Likewise, no adult insect (imago) competes with its larval stage for food.
Regardless of the species in which it is taking place, the process of metamorphosis implies several universal events:
destruction of specifically larval structures (abdominal legs of caterpillars, gills and tail of tadpoles),
adaptive remodelling of tissues that persist in the adult stage (nervous system, excretory organs),
development of structures unique to adults (wings in insects, lungs in amphibians).
In insects and other arthropods, moults occur during development and growth. Arthropods are covered with a cuticle: a hardened extracellular shell that cannot expand. The cuticle must be shed and replaced periodically to allow growth and changes in body form. At each moult, the epidermis withdraws from the old, partially dissolved cuticle and subsequently expands, forming folds and secreting a new, soft cuticle beneath the old one. When the old cuticle is sloughed off, the epidermis and the new, still elastic cuticle expand. Once the individual has hatched, it supports expansion and unfolding of epidermis and cuticle by generating hydrostatic pressure with compressed body fluid.
Metamorphosis in insects is the transformation of the larval into the imaginal (adult) phenotype. Among the various insect orders, metamorphosis takes two broadly different forms:
1.
Hemimetabolous development (Greek: hemi = half; metabole = change).
In its basic organization the larva already resembles the imago in hemimetabolous species and even has compound eyes, although the early larval body is very small and lacks structures such as wings and genitalia. With each moult, the size and appearance of the larva (often called a nymph in hemimetabolous insects) approaches the size and appearance of the final imago (Fig. 20.1e). Sometimes, only minor remodelling of the larval body is necessary (paurometabolic development); in other species conspicuous larval organs must be destroyed (such as gills in may-flies or the extensible labium of dragonflies, with its hook-like jaws). Frequently, the nymph enlarges the wing pads rather suddenly in the last two moults, but a pupa does not occur.

Fig. 20.1
(a – g) Model animals related to metamorphosis and its hormonal control. (a) Xenopus laevis, the clawed frog. (b) Axolotl (Ambystoma mexicanum); (B1) neotenic form with lifelong gills, albino; (B2) Axolotl after thyroxine-induced complete metamorphosis. (c) The hydrozoan Hydractinia echinata. Planula larva and primary polyp, founder of a new colony, arisen from the planula. Blue fluorescence marks the chitin envelope of the stolons which elongate by growth, branch like blood vessels, and bud new secondary polyps. (d) Aurelia aurita, moon jellyfish; (d1) asexually produced ephyra larva of Aurelia; (d2) Mature jellyfish. (e) Schistocerca gregaria, desert locust, example of a hemimetabolous insect with stepwise approach to the mature adult form (imago). In the sub-adult stage wings (arrow) are being formed but are not yet fully developed; (e2) The adult imago is capable of flight and sexually mature. (f) Manduca sexta, a holometabolous butterfly. (g) Araschnia levana, the map butterfly; (g1) Spring form, (g2) Summer form, also known as Araschnia prorsa, (g3) Caterpillar, (g4) Pupa, which hibernates. Pictures from teaching collection of WM, (f) Manduca sexta photos kindly provided for reproduction by Prof. Richard G Vogt, University of Washington; he is owner of the copyright
Examples of hemimetabolous development include wingless insects (Apterygota), grasshoppers (Orthoptera), cockroaches (Blattidae), termites (Isoptera), earwigs (Dermaptera), may-flies (Ephemoptera), dragonflies (Odonata), bugs (Hemiptera), sucking lice (Anoplura), and cicadas (Homoptera).
2.
Holometabolous development (Greek: holos = whole, complete). Adult structures arise in holometabolic forms from larval cell areas specified for their future task and known as imaginal discs (Figs. 20.2 and 4.41). From the egg, the young hatches with a worm-like segmented body (called the caterpillar, grub, or maggot). The larva may have short legs; it lacks wings and compound eyes. The successive larval instars increase in size through several moults but do not abandon their worm-like appearance until complete metamorphosis takes place in two major steps. In the first step, the last instar larva transforms into an immobile pupa that is encased in a protective cuticle with a form and colour characteristic of this stage. In the second step the imago is formed within the pupa cuticle. Beneath the cuticle, many larval organs break down and are absorbed by phagocytic cells, while adult structures arise concurrently. At the end of pupation, the finished imago hatches. Some of these dramatic events are described for Drosophila in more detail below.
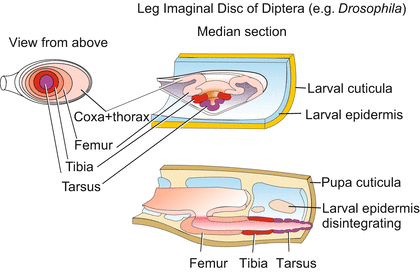
Fig. 20.2
Holometabolous development of a leg imaginal disc in the fly; its transformation in the course of metamorphosis. Colour codes indicate that distal structures are pulled out from the central area of the disc on the model of a telescope
Holometabolous development is characteristic of goldeneyes and ant lions (Neuroptera), caddis flies (Trichoptera), moths and butterflies (Lepidoptera, Fig. 20.1f, g), true flies and mosquitos (Diptera), fleas (Siphonoptera), beetles (Coleoptera), wasps, ants and bees (Homoptera).
It may be emphasized that all these phenotypic forms, embryonic stage, larva, pupa and imago derive from the same genome. At present it is inconceivable that we will ever be able to logically deduce alone from the sequence of base pairs in the DNA in which forms of appearance an organism can exist.
20.1.2 In Metamorphosis, Dramatic Remodelling Occurs Down to the Molecular Level
Metamorphosis entails reshaping and restructuring at every level of the organism, from the body’s morphology and anatomy to its physiology and cellular machinery. Such multiple changes are initiated and synchronized by hormonal signals, which are discussed in the next chapter. But as in all hormonally-controlled events, various tissues and organs respond quite differently to changing levels of hormones, depending on their programming in previous stages of development. To exemplify the remarkable range of metamorphic events, we will compare the metamorphoses of an amphibian (frog) and an insect (the fly).
Frog
Destructive and constructive metamorphic processes proceed gradually and take weeks. There is no quiescent stage; the tadpole has to manage a smooth changeover to the adult organization while it is moving and feeding.
Since the type of locomotion will change from smooth swimming to jumping with legs, hindlimbs emerge in prometamorphosis. During the climax of metamorphosis the tadpole extends its previously prepared but hidden forelimbs out of the branchial cavity, and gradually resorbs its tail (Fig. 20.3).
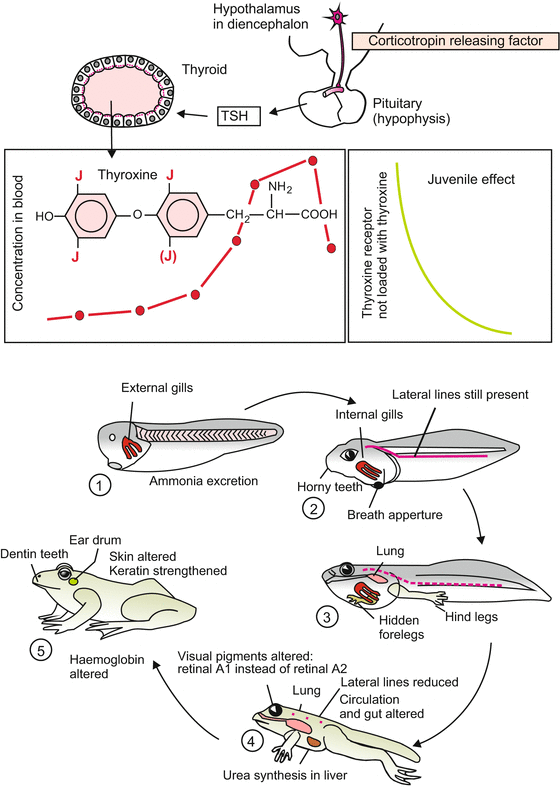
Fig. 20.3
Metamorphosis of the frog and its hormonal control (from Mueller and Frings: Tier- und Humanphysiologie, Springer Science 2009, slightly modified)
When the lungs are formed, the gills vanish. The transition in the mode of breathing is accompanied (and made feasible) by reconstruction of the circulatory system. The aortic arches and several large body vessels are restructured. In the erythroblasts a new isoform of haemoglobin, that binds oxygen less avidly, is synthesized.
To avoid desiccation, the skin is made tighter with adult keratins and interspersed with mucus glands; the eye becomes protected by an eyelid.
The horny teeth for tearing plants disappear; a long tongue develops and the intestinal tract is adjusted to a carnivorous diet.
While the tadpole can readily release ammonia into the surrounding water by diffusion (ammoniotelic excretion), in metamorphosis the liver and kidney are retooled with new sets of enzymes enabling the frog to convert ammonia into urea (ureotelic excretion).
The lateral line sensory system used by the larva to detect slow wave or current movements in water is reduced. The first branchial pocket is remodelled and becomes the ear tube (pharyngotympanic tube), closed by the tympanic membrane and incorporating the sound-transmitting columella.
In the retina of the eye the visual pigments are exchanged: the fish-type porphyropsin (opsin + retinal A2 molecule) is replaced by rhodopsin (opsin + retinal A1) characteristic of terrestrial vertebrates.
Drosophila
The larva undergoes a holometabolous (complete) metamorphosis. When entering the prepupal stage, the third and last instar wanders around seeking a place in which to transform into a pupa. Having found a suitable location, it cements itself onto the substrate with a glue produced in its salivary glands. Now the larva softens its last larval cuticle and inflates into a barrel-like shape. The expanded cuticle is re-hardened and is now called the puparium. Beneath the protective envelope most larval tissues are destroyed. The fly is constructed like a mosaic, from residual imaginal discs (Figs. 20.2 and 4.41) and imaginal cells.
The exterior epidermal structures that become covered with a cuticle are manufactured from imaginal discs. In Drosophila most of the adult is built from 10 pairs of such discs (Fig. 4.41). Unlike caterpillars of butterflies, in dipteran larvae these discs are not seen as epidermal thickenings but lie hidden inside the body. However, the discs do not originate inside the body.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


