Chapter 7 Locomotory behavior
Chapter contents
Fetal movements
The movements of the equine fetus have been studied with ultrasonography in a bid to explore the extent of normal locomotion in utero and its roles in optimizing presentation before labor and preparing the musculoskeletal system for work immediately post-partum. In addition, ultrasonography has demonstrated that sucking and swallowing is common in fetal foals, a finding that explains the development and strength of the sucking reflex at birth.1
Simple movements, such as the extension or flexion of a limb or the vertebral column, can be detected from the third month of gestation.1 As the fetus matures, these combine and become repeated to form complex movements that eventually appear coordinated.1 The hooves are tipped with collagenous eponychia, which prevents damage to the amnion and beyond. Most of the coordinated kinetic activity of the foal, including extension of the forelimbs and head toward the maternal pelvis, occurs well in advance of uterine contractions, sometimes as much as 12 days earlier.1 These final adjustments are considered critical in the avoidance of dystocia.2
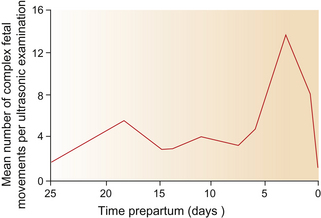
While simple movements may occur 55 times per hour from the fifth to the ninth month of gestation,1 complex movements as frequent as 84 times per hour have been recorded in the 3 days pre-partum.1 The frequency of the final complex movements that ultimately change the fetal position from supine to prone and occasionally from posterior to anterior1 is illustrated in Figure 7.1.
Infant growth and movements
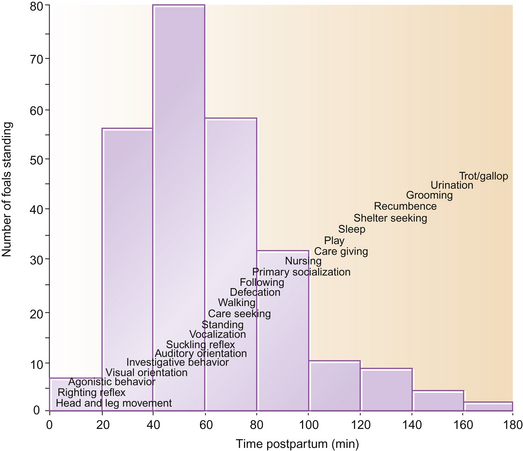
An appreciation of the normal development of foals is useful for practitioners, since deviations from the typical developmental pattern can indicate abnormal progression in a neonatal foal.3 Fraser1 suggests that five steps can be identified in the development of the primary mobility required of a foal before it takes its first mouthful of milk and the self-reinforcement of feeding can take place (Table 7.1).
Table 7.1 The steps involved in achieving primary mobility in the equine neonate
| Step | Foal’s responses | Dam’s responses |
|---|---|---|
| Recumbent coordination | ||
| Rising and quadrupedal stability | ||
| Ambulation | ||
| Maternal orientation | ||
| Teat-seeking and sucking |
After Fraser.1
The order in which features of the ethogram emerge is described in Figure 7.2. Foals that accomplish one phase quicker than the mean often show similar precociousness in other domains.1 Perhaps because they have shorter long bones and therefore require less leverage to stand, pony foals are usually stable on their feet within half an hour,1 much earlier than neonatal Thoroughbreds. The time taken to stand depends on the sex of the foal (average for Thoroughbred fillies being 56.3 minutes and for colts 70.6 minutes, n = 127)4 – a difference attributed to the lighter bodyweight of fillies, who therefore require relatively less strength to rise. Unsurprisingly, fillies are the first to feed, generally having their first drink 1.5 hours after birth while colts take an average of 2 hours before feeding.1
After the first drink, a melange of rest and sleep occupy most of the foal’s first week but, along with locomotory play, this is punctuated by feeding every 15–30 minutes.1
A daily tally of motor activities in the equine neonate is given in Table 7.2. The prevalence of vigorous stretching (or pandiculation) from the first day of life suggests that it is not only pleasurable as a comfort behavior, but also that it occurs as a result of uterine confinement.1 Stretching may occur when the foal is recumbent or upright (Fig. 7.3). The importance of stretching is dealt with elegantly elsewhere1 but can be summarized in terms of its role in athletic development, growth and joint correctness. By the third day of life, estimates of 80 or so stretches per day have been recorded.1 Some support for the putative orthopedic benefits of stretching comes from the coincidental peak in the frequency of stretching on day 3 of life, the age by which marginally contracted tendons have self-corrected. It would be interesting to investigate the effect of post-foaling thoracic trauma5 on the frequency of holistic stretching.
Table 7.2 Frequency of motor patterns in the equine neonate
| Activity | Daily frequency |
|---|---|
| Defecation | 2–4 |
| Urination | 4–10 |
| Walking | 8–14 |
| Sleeping | 20–25 |
| Sucking | 18–24 |
| Stretching (recumbent and upright) | 40–60 |
After Fraser.1
In the first 6 weeks of a foal’s life, most vigorous exercise takes the form of play,6 but the motivation to rest, play and feed changes with age. Crowell-Davis7 identifies three periods of behavioral development in foals, all of which involve different proportions of locomotory activity:
Adult kinetics
Horses have been shown to travel on open ranges (for example, to water) up to 65–80 km daily. For pastured horses, movement during grazing is the main initiator of locomotion, and an estimated daily figure for kinetic behavior in this context is 20 km.1 The distance that grazing horses travel depends on the location of water, the availability of food and the time that has to be spent foraging.
As a means of returning horses to their home range and indeed their social group, homing can occur over distances of more than 15 km.3 The stimuli used to guide this locomotion probably involve olfaction that relies on ground-based stimuli, but they remain the subject of some debate,8,9 especially since homing may even traverse water.3
While incessant walking has been reported as a presenting sign in serum hepatitis,3 route-tracing in the form of box-walking (stall-walking in the USA) is far more common and emerges as a stereotypic behavior in animals that fail to cope with social isolation (see Ch. 5). Conversely, stereotypies are reported less frequently as exercise is increased.10 Locomotion is useful as an indicator of distress, especially when conducted at the expense of feeding; for example, solitary horses walk and trot three times more often than those that can make auditory, visual and physical contacts with other horses.11 Furthermore, locomotion’s role in recreation is considerable, with 75% of the kinetic activity of foals, for example, being in the form of play.12 The need for spontaneous exercise is emphasized by reports of post-inhibitory rebound in locomotion among horses after periods of confinement.13
While locomotion is an integral part of equine anti-predator strategy and ingestive behavior, it is also used in communication and courtship. Exercise tends to affect other behaviors. For example, exercised horses spend more time drinking and lying but urinate less than unexercised controls.14 Tantalizing insights into the effect of training on behavior have been offered by studies showing that horses that lose races tend to be more aroused and require greater control in the parade ring/mounting yard than winners.15 Some of the more important dependent variables from these studies are summarized in Table 7.3. For a more comprehensive list and detailed consideration of likely reasons for these variables being significant to performance, readers are directed to an informative and entertaining book, Watching Racehorses.15
Table 7.3 Some of the features of a racehorse’s behavior and presentation that can be used to predict poor performance (not winning)
| Location | Variable | Extent to which this variable predicts poor performance |
|---|---|---|
| Birdcagea | Weaving and repetitive head movements | ******** |
| Kicking | ***** | |
| Pawing | ** | |
| Parade ring | Crossover (figure-of eight or grackle) noseband | ******** |
| Gaping mouth | **** | |
| Twisting neck | *** | |
| Nose roll (sheepskin noseband) | *** | |
| 3rd metacarpal bandages | *** | |
| Other bandages | ************** | |
| Pacifiers (meshed eye protection) | ** | |
| Mounting yard | Slow gait | ******** |
| Fast gait or circling | ***** | |
| Bucking | ***** | |
| Balking | **** | |
| Courtship behaviors | *** | |
| Ears pointing laterally or aborally | ** | |
| Defecating | ** | |
| Flicking ears | * | |
| Track | Late | ******** |
| Conflict behaviors | **** |
a In Australia, racehorses can be observed in a gallery of open-front standing stalls called the birdcage, prior to being paraded (Fig. 7.4).
Adapted from Watching Racehorses15 with kind permission of Geoffrey Hutson.

Figure 7.4 Horse in cross ties pawing in the standing stalls of a birdcage gallery at a racecourse.
(Reproduced, with permission, from Hutson.15)
Sidedness and symmetry
Although the forelegs alternate in leading during grazing, the time horses spend with the left leg advanced is generally longer than for the right (Fig. 7.5). This manifests as a significant directional bias to graze with the left foreleg in advance of the right.16 This tendency seems breed-dependent with Thoroughbreds showing it more than Standardbreds who in turn show it more than Quarterhorses.17 If advancing a forelimb when grazing reflects greater mobility on that side of midline, then it is possible that the brains of left-preferent animals are right-hemisphere dominant. However, a counter argument is that the non-advanced limb is the more critical for survival because it supports more weight, reflects greater agility on the weight-bearing side of the animal and is arguably better positioned to launch the animal into a flight response and a more dominant left turn (owing to the abduction of the right foreleg in the stance phase). We are yet to study how this affects ridden work, such as canter leads and preferred side for poll-flexion and lateral bend.

Figure 7.5 Even in the absence of lameness, some foals lock in a strong preference for an unbalanced grazing stance before weaning. There is a general population bias to advance the left foreleg. (Figure from www.avma.org)
Even in symmetrical gaits, such as the trot and the pace, asymmetries often become evident at high speeds. In pacers, temporal gait variables, including suspension times, may differ between the left and right couplets, while in trotters high speeds have been used to expose left-right asymmetries that are particularly apparent in the hindlimbs.18 This is not necessarily an effect of training to work in one direction. Slight individual tendencies toward either left or right laterality in general responses, such as in lying down,1 and in foraging, have been reported by others.19,20 Kinematic differences between the left and right limbs have also been reported in 8-month-old Standardbreds that have yet to be trained.21
It is reported that, when jumping, most horses seem to lead with and land with the right leg.1 However, other workers have shown this preference only when horses are jumping on a right-handed turn.22
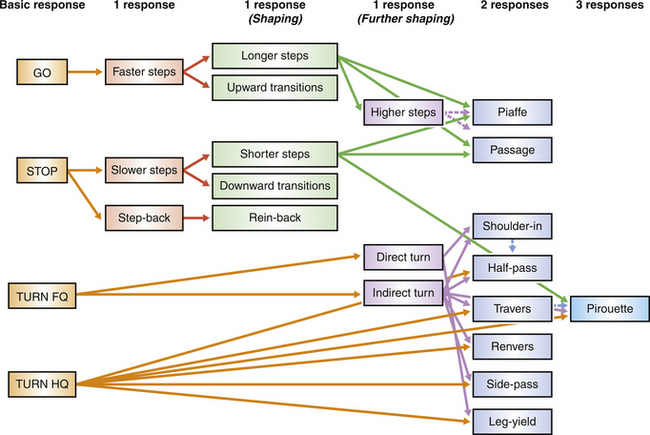
Lateral flexion
In the leg yield, the neck and vertebral column are straight except for a slight flexion at the atlanto-occipital joint that flexes the head away from the direction of motion. For the movements in which the vertebral column is flexed, it is easiest to consider the horse moving relative to an imaginary straight line. In some of the movements the neck and forequarters align with the line and the hindquarters deviate to the side (travers, renvers). In other movements the hindlegs follow the line while the forequarters deviate from it (shoulder-in, contra shoulder-in) (Fig. 7.6).
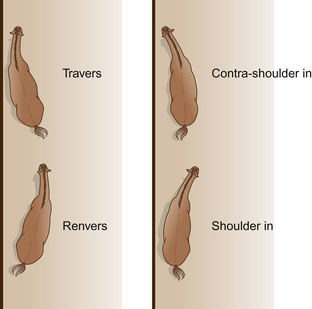
Naming lateral movements depends on the direction in which the horse’s body is bent relative to the direction of movement around the arena – clockwise (on the right rein) or counter-clockwise (on the left rein). When the convex curve of the body forms the leading edge of the horse as it travels forward, the maneuver is called shoulder-in. For the reverse maneuver, in which the direction of flexion is the direction of locomotion, the terms travers or renvers are used. Half pass (Fig. 7.7) is the same as travers except that it is performed on a diagonal line across the arena.

Simple turns and pirouettes
Turns on the forehand and turns on the quarters are achieved by the rider signaling for more movement by the quarters and forehand respectively. In their elementary forms they are conducted at the rhythm of a collected walk, but when the same principles are applied in advanced horses, elaborate balancing maneuvers, such as canter pirouettes (Fig. 7.8), result. However, despite the theoretical requirement that a good canter pirouette be conducted with the same tempo and rhythm as the collected canter, this is rarely the case. Analysis of individual medal finalists at the Barcelona Olympics showed that pirouettes were performed at about two-thirds of the tempo of a collected canter, had no suspension phase, and were dissociated to the extent that they could be considered to have a four-beat rhythm.23

This chapter will consider the gaits and maneuvers of ridden horses and those at liberty (Fig. 7.9).
Gaits
By measuring oxygen consumption in trained ponies that change gait on command, Hoyt & Taylor24 demonstrated that walking is the gait that needs least energy at low speeds, trotting at intermediate speeds, and galloping at high speeds. This accounts for the evolution of gaits as adaptations for economical locomotion within a range of speeds.
It is argued that, in many ways, the physics of equine locomotion have evolved to their limits in that, for example, if the horse’s legs were any longer, its hindlegs would be repeatedly interfering with its forelegs, as happens when the hurried (so-called butcher’s boy) trot leads to forging. Budiansky25 points out that this is the reason species such as the giraffe, with its relatively longer legs, cannot trot.
The care with which horses place their feet at speed is remarkable. With only one toe upon which to land there is little margin for error. Balance is matched by the tremendous care horses take over their hooves. This book would not be a book for hippophiles without the oft-quoted adage ‘no foot – no horse’. This appears here to remind us that horses seem to live by the very same dictum. Their motivation to avoid getting their feet trapped accounts for the reluctance of some horses to accept hoof-care. Considerable neural circuitry is required for any animal to plot a safe course for four limbs, especially as the feet disappear from sight so rapidly. To perform this at the speeds horses reach and through the terrain they can traverse helps to explain why so much of the brain has to be devoted to coordinating limbs and controlling locomotion (see Ch. 3).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


