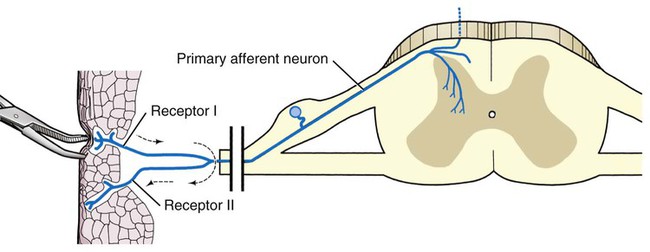
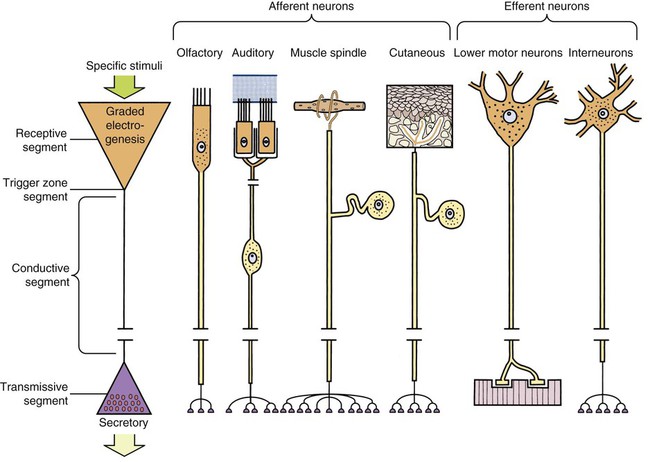
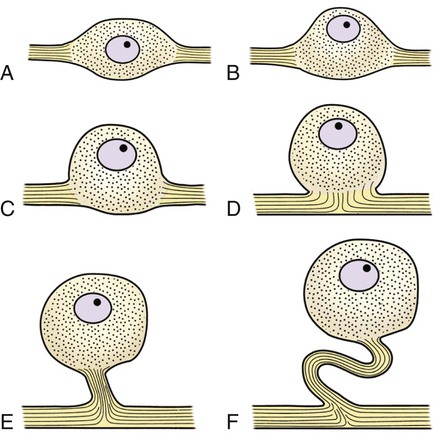
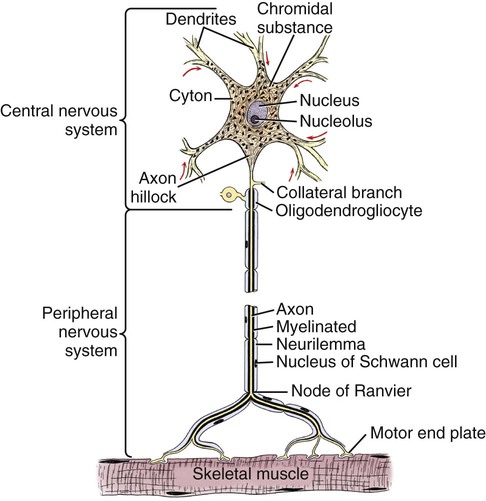
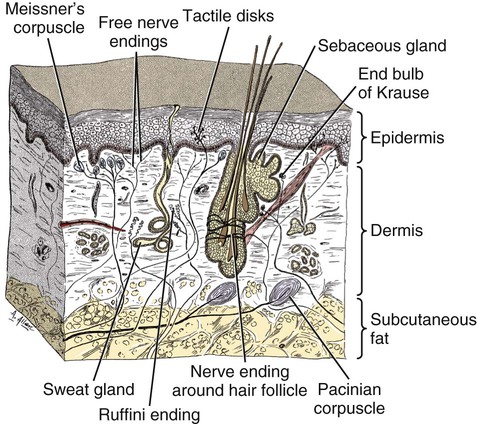
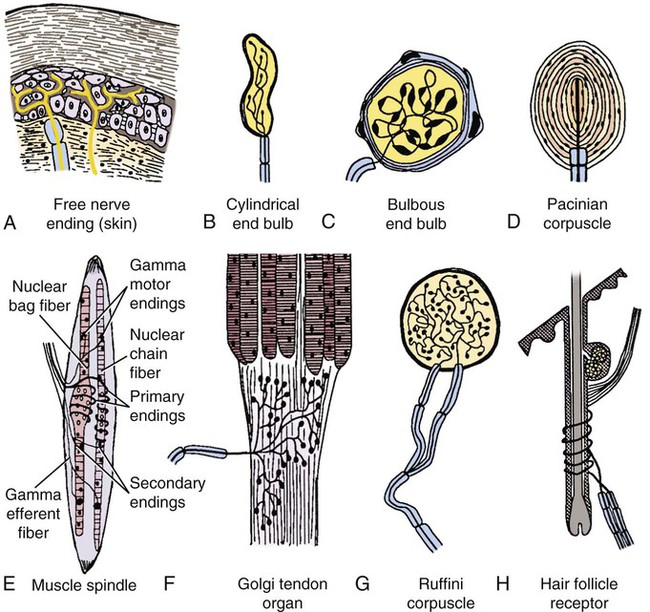
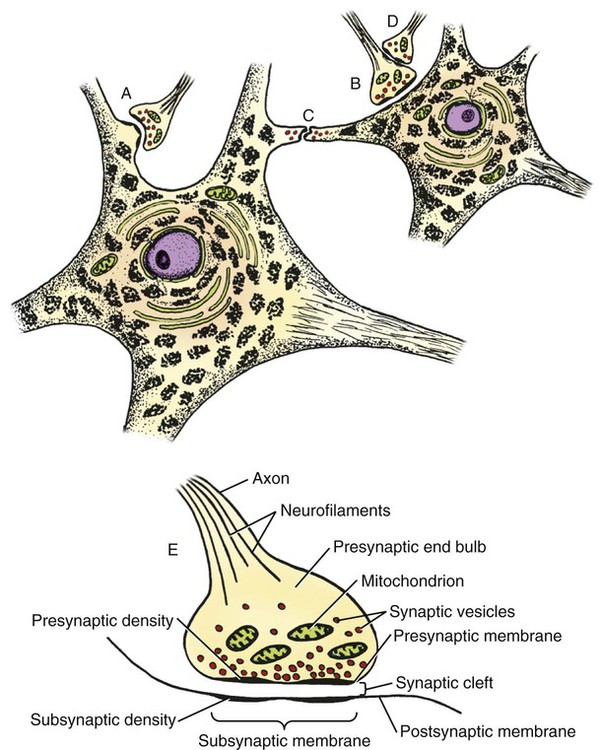
The cells constituting the nervous system are highly specialized for receiving stimuli, by means of receptors and highly specialized sense organs, for transmitting and storing information, and for initiating responses to stimuli (Fig. 14-1). Cells composing the nervous system are of two types: neurons (neurona) that receive and transmit impulses; and supporting cells that regulate the rate of axonal conduction of impulses, hold the neurons in place, and prevent the neuronal impulses from spreading between neurons that are not linked by synapses. Pseudounipolar cells embryologically start out as bipolar cells, but the cell body grows out on one side, and the two processes emerge from a tortuous neck of the cell (Fig. 14-2). Both processes are structurally axons. Bipolar cells have two processes, classified structurally as axons. Multipolar cells have many processes, one axon and the rest classified as dendrites (Figs. 14-1 and 14-3). Olfactory neurons are unipolar, auditory neurons are bipolar, muscle spindles and cutaneous afferents are pseudounipolar, and lower motor neurons (LMNs) and interneurons are multipolar. The location of the cell body is not critical to the functioning of the various neurons. The cell body may be in the receptive segment such as olfactory afferent neurons, LMNs, and interneurons. The cell body may be in the conductive segment such as auditory and vestibular afferent neurons or the cell body may branch off the conductive segment such as all other afferent neurons. The transmission segments end on other neurons or on muscle or gland cells in the case of motor neurons. The endoplasmic reticulum (reticulum endoplasmicum), or cytoplasmic chromidial substance, of neurons is concentrated in the cell body and in parts of dendrites close to the cell body (see Fig. 14-3). The endoplasmic reticulum is sparse in one region of the cell body, called the axon hillock (colliculus axonis), where the axon arises, and in the axon and in the distal branches of dendrites. Because the endoplasmic reticulum of the cell body is so densely packed and is highly organized, it appears in microscopic sections as basophilic clusters, called Nissl substance (substantia chromatophilia) or Nissl bodies. The Nissl substance stains blue with basic dyes, such as thionin, cresyl violet, toluidine blue, and methylene blue. Based on both structure and function, every neuron can be divided into five segments (see Fig. 14-1): cell body, receptive segment, trigger zone segment, conductile segment, and transmissive segment. The receptive segment of the neuron, also referred to as the dendritic zone, varies in shape and location. It is the site on a neuron where a stimulus results in a change in the cell membrane potential that will generate a nerve impulse in the trigger zone. In primary afferent neurons the receptive segment is in the receptor (the ending in a sense organ). In the olfactory system, the receptive segment is the distal end of the cell body of the neuron that is acted on by the chemical odor. In interneurons and LMNs the dendrites and cell bodies are the receptive segment and are acted on by chemical neurotransmitters released by synaptic terminals of other neurons. In some instances, the receptive segments may be on the synaptic end bulbs (the transmissive segment) of a neuron (axoaxonic synapses; see Fig. 14-7D). Receptive segments of neurons function by developing a nonpropagated change in the membrane potential of the neuron that is spread decrementally along the neuron to affect the trigger zone segment of the neuron (see Fig. 14-1). The conductile segment of the neuron is commonly referred to as the neuronal fiber or axon. In the case of bipolar, or pseudounipolar, neurons, the conductile segment is divided into two parts, depending on the relationship of the segment to the cell body. The conductile segment of a primary afferent neuron that begins at a receptor in the periphery is referred to as the peripheral conductile segment. (As stated earlier, in many histology textbooks this is erroneously called a dendrite.) The conductile segment of these cells that carries the nerve impulses away from the cell bodies and into (and within) the CNS is called the central conductile segment. The conductile segments of olfactory neurons, interneurons, and LMNs begin at a specialized part of the cell body referred to as the axon hillock, and the conductile segments, the axons, extend away from the cell bodies to their termination at the transmissive segments, which are the location of the synapses with other neurons or effector organs. In alpha, beta, and gamma LMNs, the axon begins at the axon hillock and has a long peripheral conductile segment within the PNS that conducts nerve impulses to the transmissive segment. Spinal cord autonomic lower motor neurons have preganglionic axons that have synapses with ganglionic autonomic neurons (postganglionic) (see Fig. 14-11). Ganglionic autonomic neurons have postganglionic axons that activate smooth or cardiac muscle or glands or, in the case of the adrenal medulla, release the neurotransmitter directly into the blood stream. The conductile segments of all neurons are capable of conducting impulses in either direction, depending on the site on the neuron where the neuronal impulses are generated (see Fig. 14-4). In some primary afferent neurons that have branching peripheral conductile segments, each with a receptor associated with them, a nerve impulse generated at the trigger zone of one branch can travel toward the CNS via the conductile segment, then travel away from the CNS via another branch of the conductile segment. Those nerve impulses traveling away from the trigger zone are referred to as being conducted orthodromically; those traveling away from the CNS are referred to as being conducted antidromically (in human primary afferents, anterograde). Dendrites of interneurons and LMNs are not included in the conductile segments of these neurons because dendrites of these neurons do not conduct nerve impulses, but the bioelectrical change initiated in them by synapses spreads decrementally (not regenerated as a nerve impulse) to the cell body (orthodromically) or away from the cell body (antidromically). The transmissive segment of a neuron is located at the terminal end of the conductile segment, the axon, called the telodendron of the neuron. The transmissive segment of primary afferent neurons, interneurons, lower motor neurons and projection neurons of the CNS is modified to form a synaptic end bulb (see Fig. 14-7), where neurotransmitters are released on the receptive segments of interneurons or efferent neurons. The transmissive segments of alpha, beta, and gamma motor neurons are modified to form neuroeffector junctions (motor end plates), where neurotransmitters are released to act on the effector (muscle fibers). The peripheral terminal ends of sensory (afferent) axons are modified to form specialized structures, called receptors (receptori) (Figs. 14-5 and 14-6). These may, or may not, have connective tissue capsules (corpusculi nervosi capsulata) surrounding them. Receptors are specialized to generate neuronal impulses in their axons when a specific stimulus or sensory modality is applied to them. These include chemical, mechanical, thermal, or electromagnetic energy. In humans, each receptor type has a low threshold for a specific modality thus is most sensitive to that modality. Free nerve endings (terminationes nervi libera) (see Fig. 14-6A) are responsive to warm or cold temperatures, noxious stimuli that are potentially or actually damaging to tissue and may be associated with perception that is exhibited as pain, and crude mechanical stimulation. End bulbs of Krause (corpora bulboidei) (Fig. 14-6B and C) respond to slow movement (flutter). Pacinian corpuscles (corpusculi nervosi capsulata) (Fig. 14-6D) respond to rapid vibration. Muscle spindles (fusi neuromusculari) (Fig. 14-6E) detect muscle length and the velocity at which a muscle is being stretched. Golgi tendon organs (fusi neurotendini) (Fig. 14-6F) detect muscle tension. Ruffini corpuscles (corpora tacti) (Fig. 14-6G) respond to tangential stretching. Hair follicle receptors (terminationes folliculi pili) (Fig. 14-6H) respond to movement but not to continual pressure, and thus are referred to as mechanical motion detectors. Merkel discs (epithelioidocyti tacti) (Fig. 14-5) and tactile corpuscles (toruli tactiles) (Fig. 14-5) respond continuously to sustained perpendicular pressure. Other afferent as well as efferent axons may have specialized branches (telodendria) at their terminal endings. These telodendria end by forming a synaptic end bulb (synapsis vesicularis), in which synaptic vesicles (vesicula synaptica) containing a neurotransmitter are concentrated (Fig. 14-7). The synaptic end bulbs contact another neuron (dendrites, cell body, or occasionally an axon). Efferent axons ending on muscle form motor end plates (terminationes neuromusculi). The telodendria form synapses where they release neurotransmitters that affect the membrane channels of the other neuron or the muscle fiber.
Introduction to the Nervous System
General
Structure of Neurons
Functional Segments of Neurons
Receptive Segment
Conductile Segment
Transmissive Segment
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Introduction to the Nervous System
Only gold members can continue reading. Log In or Register to continue
