Chapter 14 Gastrointestinal Physiology and Nutrition
The rabbit is a monogastric, hindgut-fermenting herbivore with a complex and unique digestive physiology. Given the wild rabbit’s ecologic role as a small prey species, its digestive strategy has evolved to permit efficient digestion of fibrous vegetation without the need to store large volumes of food within the body. The gastrointestinal tract features a simple stomach, a well-developed cecum, and a particle-dependent separation mechanism within the proximal colon. Fine particles and solutes are selectively retained as substrates for microbial fermentation within the cecum, while indigestible fiber is rapidly expelled.50 The production and ingestion of cecotrophs (packets of cecal contents) contributes significantly to the rabbit’s digestive efficiency.
Rabbit Gastrointestinal Physiology
Ingestion of Food
Rabbits are described as selective feeders; in feeding on natural vegetation, they select the most tender, succulent plant parts (i.e., the parts that are most nutrient-dense and lowest in available cell walls).12 Their natural diet consists of the preferred succulent buds and young leaves of bushes, and they routinely graze on grasses, weeds, and even the bark around bushes and trees.7 They have a relatively high metabolic rate and a fast feed-transit time (19 hours)32; moreover, the practice of selective feeding (often termed “concentrate selection”) allows them to meet their dietary requirements while minimizing the volume of food that must be eaten. Rabbits eat approximately 30 times per day (2-8 g of food per time) over 4- to 6-minute periods.48 The selection of food is based on olfactory cues and tactile information obtained via the sensitive vibrissae around the nose and lips.27 The incisors have evolved to cut through vegetation with a vertical slicing motion, while the cheek teeth are responsible for grinding the food before it is swallowed. Jaw movements during mastication are reported to be up to 120 per minute5 and amylase-containing saliva is secreted continuously during the process. Jaw movements feature a lateral motion, which helps keep the constantly growing teeth worn down to maintain effective occlusal surfaces. Hunger is stimulated by a dry mouth and gastric contractions or a decrease in blood levels of metabolites such as glucose, amino acids, volatile fatty acids, or lactic acid.18
Stomach
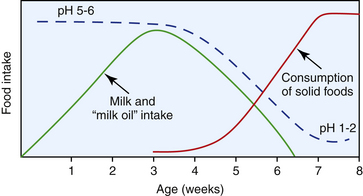
The gastric pH of the rabbit varies diurnally but is generally very acidic compared with that of other species; postprandially it can drop to as low as 1.0 to 2.0, whereas following the ingestion of cecotrophs it rises to 3.0.4 This low pH effectively sterilizes ingesta. Juvenile (preweaned) rabbits have a much higher gastric pH (5.0-6.5) (Fig. 14-1), which promotes the survival and passage of ingested bacteria, facilitating the establishment of the vitally important large intestinal flora.27
The stomach usually contains hair, food, and fluid even after 24 hours of fasting or anorexia. Gastric transit time is approximately 3 to 6 hours.10
Small Intestine
The small intestine of the rabbit is the primary site of absorption of many nutrients, such as amino acids, lipids, monosaccharides, and electrolytes. A wild-type diet, however, contains very few of these nutrients in a form suitable for absorption as the ingesta pass from the stomach into the small intestine. From a nutritional perspective, vegetation is predominantly composed of lignin, cellulose, hemicelluloses, oligosaccharides, pectins, and plant proteins bound to cell wall constituents, and these pass through the small intestine largely unaltered. Conversely, nutrients present within ingested cecotrophs are available in a form that does permit absorption across the small intestinal epithelium. Transit time through the small intestine is fast: 10 to 20 minutes through the jejunum and 30 to 60 minutes through the ileum,10 which limits the time available for absorption at this site.
Bicarbonate is secreted into the proximal small intestine and functions to neutralize the acidic digesta leaving the stomach. The bicarbonate is then absorbed in the jejunum. Unlike the situation in many other mammals, pancreatic amylase is not a significant contributor to digestion.27
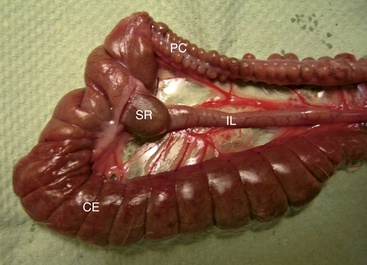
To complement its role in the absorption of nutrients, the small intestine appears to have an important immune function. There are extensive aggregates of lymphoid tissue (Peyer’s patches) throughout the intestinal tract of the rabbit, most prominently in the distal jejunum. Additionally, the terminal ileum enlarges into a dilation known as the sacculus rotundus at the ileocecocolic junction (Fig. 14-2). The sacculus rotundus (also known as the ampulla ilei or ileocecal tonsil) is unique to the rabbit and is made up primarily of lymphoid tissue.
Large Intestine
The colon is functionally divided into two parts: the proximal colon, which is grossly identified by the presence of distinct haustra (sacculations), and the distal part, which is devoid of haustra. The proximal colon is the site of separation of ingesta into the digestible and indigestible fractions. The separation process is facilitated by a combination of functional anatomy and colonic motility. The digestible particles (which tend to be shorter) settle near the mucosa and are subsequently propelled in a retrograde direction back into the cecum via a series of coordinated contractions. At the same time, the longer, coarse, indigestible particles tend to remain in the center of the colonic lumen and are directed aborally to form hard fecal pellets (Fig. 14-3).29 The indigestible fraction (sometimes referred to as the “scratch factor”) is not broken down structurally and therefore does not contribute directly to the acquisition of energy or nutrients; nevertheless, it is absolutely crucial for the stimulating normal gastrointestinal motility and for maintaining the vital physiologic processes of cell regeneration, secretion, digestion, absorption, peristalsis, and excretion.
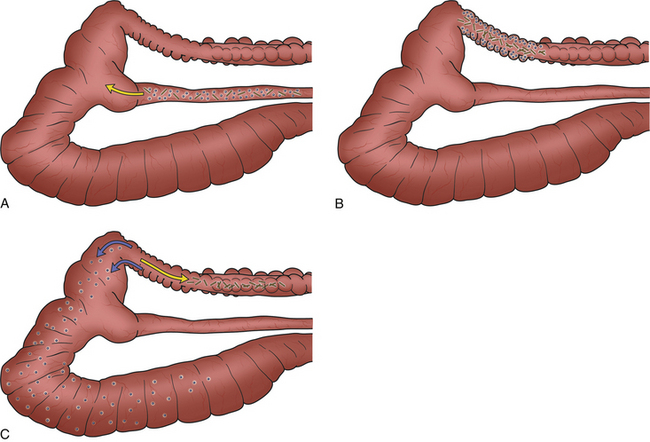
At the junction between the proximal and distal colon there is an area of thickened circular muscle that is densely innervated and vascularized. This structure is termed the fusus coli and is unique to lagomorphs. It acts as an “intestinal pacemaker,” controlling segmental, peristaltic, and haustral colonic motility and is responsible for mechanically squeezing water and electrolytes from hard feces prior to their expulsion. Reabsorption of water and solutes removed in this fashion occurs in the distal colon. The fusus coli (and hence intestinal motility) is influenced by hormones such as aldosterone and prostaglandins.27
The cecum is the largest organ in the abdominal cavity and has 10 times the capacity of the stomach. It usually contains approximately 40% of the intestinal contents. It is very thin-walled and coiled, ending in the blind-ended vermiform appendix.41 The appendix is rich in lymphoid tissue and also has a secretory function (water and bicarbonate). The cecum receives the short particles and fluid selectively retained by the proximal colon; therefore its contents are generally semifluid in consistency. Microbial fermentation is the primary mechanism by which nutrients are released from ingested food, and the retained particles directed from the proximal colon to the cecum provide the substrate for the indigenous population of cecal microorganisms. Some of the products of fermentation are absorbed directly through the cecal wall, while many others are expelled and reingested as cecotrophs.27 The nutrients present within cecotrophs are made available to the rabbit when they pass through the stomach into the small intestine.
Excretion of feces follows a marked circadian rhythm, alternating between the hard-feces phase (which coincides with feeding activity) and the cecotroph phase (which usually coincides with a period of rest at least 4 hours after feeding); this rhythm is influenced by diet, age, and reproductive status.10 Like the small intestine, the large intestine features significant amounts of gut-associated lymphoid tissue (GALT). In fact, the large intestinal GALT accounts for over 50% of total lymphoid tissue in the rabbit.46
Hindgut Flora and Fermentation
To facilitate the digestion of selectively retained materials, the rabbit’s cecum has a well-established autochthonous population of microorganisms. These microorganisms produce the volatile fatty acids acetate, butyrate, and propionate,23 which provide up to 40% of the rabbit’s maintenance energy requirement.38 The proportion of the three volatile fatty acids varies according to the time of day, the diet, and the rabbit’s developmental stage. The microbiology and ecology of the rabbit gastrointestinal tract has not been as extensively studied as that of ruminant species or humans,52 but it is widely agreed that across all mammalian species a healthy intestinal flora is vital for systemic health. The indigenous intestinal microflora is essential for digestion but also serves a protective role in relation to potential pathogens. Studies to date have identified an extensive list of organisms that may be present, including a variety of anaerobes,52 gram-positive and gram-negative facultative anaerobes,53 large ciliated protozoa, and a rabbit-specific ascosporogenous yeast (Cyniclomyces guttulatulus).20 It is generally agreed that the strict anaerobes of the genus Bacteroides are the predominant organisms within the cecum, accounting for 109 to 1010 per gram out of a total bacterial load of 1010 to 1012 per gram of cecal contents.45 Coliform bacteria and Clostridium species are occasionally isolated from normobiotic rabbits; if they are present, however, they represent a very small percentage of the total bacterial population.52 In contrast with other mammals, rabbits have very rarely been found to harbor lactobacilli.35,55 Energy is the limiting factor for the cecal microbial population36 and, as is the case in other species, the composition of the microflora in any one individual does not remain constant. The cecal production of carbohydrates and nitrogenous substances, along with cecal goblet cell mucin production, supports abundant microbial synthesis.7 The nitrogen sources for cecal bacteria are predominantly ammonia, urea, and biuret.
Disruption of the normal balance of microflora in the gut, usually with overgrowth of known or potential pathogens, is termed dysbiosis and is a common and serious clinical problem in rabbit medicine.11,53 Such alterations in normal homeostatic mechanisms within the gut may occur secondary to inappropriate therapeutic antibiotic administration, exposure to pathogenic organisms or toxins, increased glucocorticoid levels (iatrogenic or secondary to stress), gastrointestinal hypomotility, and poor dietary composition (low fiber, high carbohydrate, and high protein levels) (see Chapter 15).
Suckling rabbits are unique among nursing neonates in that they feed for only 3 to 4 minutes in every 24 hour period30 and in that they produce an antimicrobial fatty acid referred to as “milk oil.” This oil results from an enzymatic reaction that occurs in the stomach following the ingestion of doe’s milk, and it appears to be a factor in the control of the gastrointestinal microbial content of young rabbits.16 Rabbits fed milk from other species do not develop this antimicrobial factor and are more susceptible to infection.27 There is virtually no microflora in the rabbit gastrointestinal tract at 3 days of age, and although some bacteria can be found in the small and large intestines over the following 3 weeks, the stomach remains largely devoid of microflora during this time. The production of milk oil wanes as the weaning process progresses and solids replace milk.16 In this transitional period, as the production of stomach oil ceases but before gastric pH has decreased to adult levels, populations of bacteria pass from the stomach to colonize the small intestine, cecum, and colon.
Cecotrophy
Rabbits produce two types of feces over the course of a day: hard feces and cecotrophs (also termed “soft feces” or “night feces”) (Fig. 14-4).53 These two excretory products differ markedly in composition,29 and in a healthy rabbit the excretion of only one type occurs at any one time. Hard fecal pellets are composed of compressed indigestible fiber (dry matter, 52.7%) that is separated from the remainder of the ingesta in the proximal colon, whereas cecotrophs (dry matter, 38.6%) contain semiliquid cecal contents and are rich in essential amino acids, volatile fatty acids, enzymes such as amylase and lysozyme, vitamins B and K, and microorganisms including bacteria, yeasts, and protozoa.7,27,50 The protein content of cecotrophs varies between 24.4% and 37.8%, of which 81% is in the form of bacterial cells.25 Cecotrophs are excreted according to a complex circadian pattern, which opposes that of voluntary food intake and hard feces production.10 Initiation of cecotroph formation occurs when segmental and haustral contractions of the large intestine are replaced by mass peristaltic activity resulting in the expulsion of material from the cecum.15 The expelled cecal content is then packaged and covered by a mucous envelope in the colon and passed as soft pellets approximately 5 mm in diameter and arranged in clusters. The transit time for cecotrophs through the colon is 1.5 to 2.5 times faster than that for hard feces.19 They are consumed directly from the anus, a practice referred to as cecotrophy (Fig. 14-5), and are swallowed without mastication.11 The ingestion of cecotrophs is triggered by stimulation of rectal mechanoreceptors, the perception of the specific odor of cecotrophs, and blood concentrations of various metabolites and hormones.18 The percentage of cecotrophs eaten varies depending on feeding regime and dietary composition. For example, fewer cecotrophs are consumed on a high-protein diet compared with a high-fiber one.


Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


