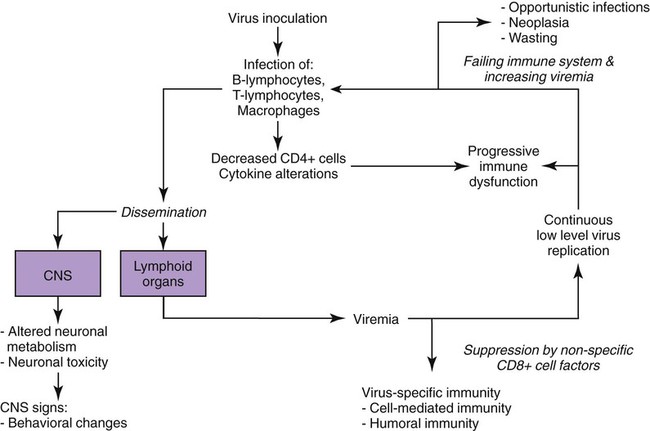
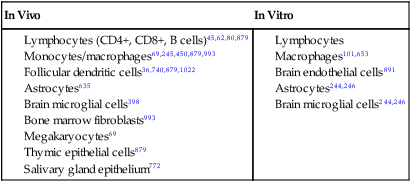
Feline immunodeficiency virus (FIV), a lentivirus that shares many properties characteristic of other lentiviruses such as human immunodeficiency virus (HIV), retains great interest as a model of lentiviral pathogenesis and prevention. Given the enormous volume of literature generated since the first description of FIV in 1986, the principal goal of this chapter is to familiarize the reader with current, clinically important concepts. Readers interested in a review of the genetic organization, biology, and life cycle of FIV, details of FIV gene function and gene products, and comparisons to other lentiviruses are referred to other sources.* Although discussion of the FIV genome and its products is not the focus of this chapter, several FIV genes have clinically important aspects. Regions in the viral integrase enzyme determine the site of binding and integration of FIV provirus into the host cell DNA, which can influence host cell function.927,928 The envelope (env) gene and its proteins768 also are of clinical importance. Field isolates of FIV are divided into several subtypes (clades) based, in part, on sequence differences in a hypervariable region of the env gene.766,1056 Within regions, numerous subtypes exist, owing to a high level of genetic recombination, especially of the env gene. Worldwide, five major subtypes have been recognized: subtypes A, B, C, D, and E. Additionally, new subtypes have been described in cats from Texas (subtype F), New Zealand, and Portugal.251,394,728,779,1071 Available studies suggest that subtypes A and B predominate in the United States and Canada, with some cats infected with subtypes C and F.37,78,850,1070a,1099 These studies also suggest regional differences in subtype distribution.1070a In Australia, the presence of subtypes A and B has been described,467,505 and in New Zealand, subtypes A and C.393a,394,503,506 In Africa, subtype A and in South America, subtypes B and E have been found.155,654,654 Subtypes B, C, and D predominate in Japan and other Asian countries, although subtypes A and E have also been observed.413,542,722,728,1032 European cats are infected with subtypes A, B, C, and D, with subtype A being the major subtype in the northern countries (e.g., Germany, The Netherlands), and subtype B being more important in southern countries (e.g., Italy).250,803,980,979 Analysis of European FIV subtypes has suggested that subgroupings within a given subtype are also possible, reflecting the genetic plasticity of FIV.980 Differences in env antigenic determinants presents potential obstacles in the development of FIV vaccines protective against widely prevalent, and different, isolates of FIV409,936,936 (see Prevention). Naturally infected cats can harbor multiple subtypes,545,748 and superinfection indicates a lack of cross-protection between subtypes.151,545,545 Evidence suggests that exchange of gene segments encoding the env protein from different subtypes can occur between isolates in superinfected cats.151 Such recombination events could be one factor in the emergence of new subtypes. Env properties are also clinically relevant because they determine cell tropism* and influence pathogenicity.480,484,531,766,1056 Interactions of FIV env proteins with host cell are critical initial steps during host cell infection, making them potential targets for therapeutic intervention.426,691,691 Env proteins are targets of immune responses,† and differences in, or conservation of, env sequences may reflect selection pressures exerted by the immune response of the infected cat. FIV env sequences evolve through acquisition of mutations during the course of infection, potentially contributing to variants that resist neutralization or contribute to disease progression.538 FIV is common worldwide, and its prevalence varies among geographic locations. Across the United States and Canada, the reported antibody prevalence of FIV ranges from approximately 4% to 24%587,613,614,629,847a; the largest North American study of more than 18,000 cats identified an overall antibody prevalence of 2.5% with an 18.2% antibody prevalence in sick cats that were tested.594 In this study, the antibody prevalence in western-region cats was lower than those in other regions, in contrast to older studies of fewer animals that documented little in the way of regional differences.167,567 In Europe, antibody prevalence is highly variable. Some countries (e.g., northern European countries)352 report few infected cats, and others, such as Italy, with large populations of free-roaming cats have prevalence rates that can approach 30%.‡ For similar reasons, Japan also has very high infection rates.721 Pockets of high prevalence in countries with low overall infection rates, such as 47% reported in one group of feral cats in the United Kingdom, can also be seen,151 likely reflecting local cat population dynamics.29 Within a given population, the prevalence of FIV in healthy cats is usually lower than in sick cats.§ Antibody prevalence in virtually all surveys is higher in male cats than it is in female cats, which is considered the result of higher rates of virus transmission among biting and fighting cats.‖ Similarly, the risk of infection is higher in cats that spend more time outdoors.352,594 Adult cats are infected more often than adolescent cats and kittens are,42,352,594,712 which again likely reflects aggressive behavior between cats as the predominant means of natural transmission. In antibody surveys, it is uncommon for enzyme-linked immunosorbent assay (ELISA)-positive results to be confirmed by other methods; thus, true infection prevalence may be overestimated, especially in the healthy cat population, because of false-positive results or potentially vaccinated cats. Evidence from retrospective antibody surveys suggests that FIV has been present in the domestic feline population since at least 1966.921 Infection with lentiviruses related to FIV has been reported in Florida panthers (Puma concolor coryi) and many other nondomestic feline species in United States zoos, as well as in free-roaming nondomestic felids in the United States, Europe, Africa, Saudi Arabia, and Asia.¶ Interestingly, presence of antibodies against FIV has been reported in spotted hyenas (Crocuta crocuta),375 but whether this observation reflects cross-reactivity with a novel hyena lentivirus or reactions arising from their exposure to lion (Panthera leo) lentiviruses is not known. The greater diversity of viral nucleic acid sequences and the decreased pathogenicity of the nondomestic felid isolates, compared with those that affect domestic cats suggest that nondomestic felids have been living with the virus longer and that the domestic cat strains may have emerged from nondomestic strains.129,750 Readers interested in learning more about lentiviral infections in nondomestic cats are referred to other sources (Web Table 12-1). WEB TABLE 12-1 Summary of Literature Addressing Lentiviral Infection in Nondomestic Felids FIV, Feline immunodeficiency virus. Domestic cats are susceptible to persistent infection with isolates from nondomestic felids, but the clinical and immunologic abnormalities that develop after infection with domestic cat isolates are typically not observed.1004,1051 Furthermore, cross-infection studies suggest that infection with a nondomestic feline lentivirus (lion or puma lentivirus) may blunt the immunologic and virologic responses to subsequent FIV infection.1048,1049 Probable transmission of an FIV isolate from domestic cats to an exotic cat species has also been documentated.729 In natural settings, FIV is transmitted primarily by parenteral inoculation of virus present in saliva or blood, presumably by bite and fight wounds, accounting for the higher prevalence in male cats. Evidence supporting the importance of this route of transmission is the observation that FIV can be found in salivary gland epithelium662,772 during acute infection, as well as in saliva, blood lymphocytes, and plasma or serum.662 Experimentally induced bites can transmit virus from infected to naïve cats. Experimentally, FIV is easily transmitted by all parenteral routes (intravenous, subcutaneous, intramuscular, intraperitoneal) using cell-free or cell-associated virus. In experimental settings, high rates (over 50%) of transmission in utero and postparturition via milk have been documented in queens with acute and chronic FIV infections.* The presence of higher viral loads in milk than in milk-secreting cells or plasma suggests that virus is concentrated in milk.9 In a given litter, some kittens can acquire infection in utero, and others will not.878 Experimental transmission has also been reported after oral,698,915 intrarectal, and intravaginal inoculation with cell-free or cell-associated virus, and these FIV mucosal transmission models are commonly used to better understand HIV biology and pathogenesis.† The feline female reproductive tract contains CD4+ and CD8+ T cells, B cells, macrophages, and dendritic cells, all known targets of FIV infection. Systemic spread following mucosal routes of inoculation can occur within days.138,740 FIV infection in cats has also been used as a model of fetal/neonatal HIV infection.98,152,152 Despite experimental evidence of FIV transmission via mucosal routes, no evidence exists for this route as being important in maintaining natural infections. Transmission from mother to kittens, in utero or postparturition, is considered a rare event under natural circumstances. Available epidemiologic and antibody surveys, however, do not exclude the possibility of occasional transmission by these routes. High mortality in FIV-positive neonates or rapidly progressive infections, as observed in some experimental studies,‡ may lead to underestimates of in utero and neonatal transmission in natural settings. Additionally, the observation that kittens born to FIV-infected mothers can have FIV provirus in their tissues, but not necessarily their blood, in the absence of detectable antibodies further complicates the understanding of congenital transmission under natural conditions.8 Infectious virus has been documented in both cell-free and cell-associated fractions of semen from acutely and chronically infected male cats.486,490 Infection can be established after laparoscopic insemination of queens with semen from infected male cats.487,488 Although the contribution of seminal transmission to natural infections is unknown, it is likely low. Horizontal transmission of FIV in multiple-cat households usually is an infrequent event, with some studies suggesting it rarely occurs, but others suggest that horizontal transmission may be common.3,780 The discrepancy could reflect behavioral differences, such as fighting tendencies, between cats in these households. Cats with positive test results for FIV DNA, but negative test results for FIV-specific antibody (so-called latent infection), have been detected in situations in which the cats with positive DNA test results had been housed in experimental colonies for long periods (months to years) with cats having positive FIV-specific antibody test results (so-called active or viral-productive infection). Interestingly, cats with latent infection were asymptomatic and did not develop typical immunologic abnormalities observed in their cohabitating actively infected cats.193 Similar cases of clinically healthy cats with latent infections have been observed in other conditions.8,739 The clinical consequences, if any, of this “latent” type of infection remain unknown at present. Experimentally, other miscellaneous modes of transmission, such as using suture contaminated with blood from an FIV-positive cat, have been documented.249 Experimental inoculation of proviral DNA, without complete virus particles, has also produced infection.796,868,868 Although infection has been established with these modes, natural infections are unlikely to occur through these routes. The pathogenesis of FIV infection reflects an interplay of a large number of factors including the age of the animal at the time of infection (young animals develop clinical signs sooner), properties of the FIV isolate (some isolates are inherently more pathogenic than others), the amount of virus used for infection, the route of infection (parenteral versus mucosal or other route), and whether the inoculum is in the form of cell-free or cell-associated (i.e., infected cells) virus.* These factors come together to affect differences in viral kinetics, the character of immune responses to FIV after infection, clinical features, and progression of FIV infection. After experimental inoculation, viral particles are cleared by tissues rich in macrophages, and viral replication then occurs in target cells of lymphoid organs (thymus, spleen, lymph nodes), and other tissues rich in lymphocytes.† Using polymerase chain reaction (PCR) or viral culture, virus is easily detected in plasma or peripheral blood lymphocytes by two weeks postinfection or even earlier662 with viremia peaking within several weeks postinoculation.67,201 FIV also spreads to mononuclear cells (lymphocytes and macrophages) in organs such as the bone marrow, lung, intestinal tract, brain, and kidney.67,879,879 Results of in vitro studies indicate that FIV-infected dendritic cells can directly transmit virus to CD4+ cells,319,971 supporting earlier studies that suggested virus-infected follicular dendritic cells in lymph nodes may infect naïve CD4+ cells migrating through the lymph node.36,1022 A burst of viral replication after activation of CD4+ cells, seen in one study, raises the possibility that FIV takes advantage of dendritic/CD4+ cell interactions to enhance infection.971 After peak viremia, circulating virus decreases to low levels as the host mounts an immune response to FIV. A vigorous, but ultimately ineffective, humoral immune response is mounted against the virus (Fig. 12-1). Generally, anti-FIV antibodies first become detectable in experimentally infected cats 2 to 4 weeks postinoculation, although exposure to lesser amounts of virus may delay the appearance of detectable responses.71 Antibodies are produced against many FIV proteins,264,286,431,870 especially those of the viral envelope, capsid, and transmembrane proteins.658 Virus-neutralizing antibodies can be detected with in vitro assays,40,131,760,1023 but neutralizing antibodies do not efficiently enter cells and therefore do not eliminate virus infection; their role in the suppression of FIV viremia in vivo is not clear.212,341,460,663,667 Some evidence suggests that the humoral immune response to the virus replicating in infected cats actually is responsible for driving the emergence of FIV variants’ resistant to in vitro neutralization.346 Interestingly, some cats have provirus- (DNA-) positive test results but have FIV antibody–negative test results (see the earlier discussion of latency, under Transmission), suggesting a mechanism by which nonreplicating virus can evade host defense mechanisms. Although a humoral immune response to FIV is documented, evidence suggests that CD8+ cells play a more important role in suppression of virus production after initial infection.123,124,179,332,414 Suppression of virus production has been demonstrated in vitro through mechanisms involving both cell-cell interactions and secretion of soluble factors, including interleukin (IL)-16 or others.579,598,598 The antiviral activity of soluble factors is not restricted to a specific FIV isolate,162,179,302,416,579 and the mechanism behind its nonspecific, noncytolytic, virus-suppressive activity may be inhibition of viral messenger RNA (mRNA) transcription.412,418 Inhibitory activity can be detected in some cats approximately 1 week postinfection and before detection of a humoral immune response.303 CD8+ cell-mediated suppressive activity is maintained through the asymptomatic period, but as infection progresses into the chronic phase, suppressor activity may wane in infected cats.302,414,414 Virus-specific cytotoxic T-cell activity emerges in the weeks after infection and also plays a role in the host’s control of virus.132 Interestingly, one study described loss of cytotoxic T-cell activity in thymectomized FIV-infected cats, but no alteration in plasma virus loads during the course of the study.389 After acute infection and suppression of viremia, cats enter a clinically asymptomatic period of variable duration. This period is not one of true viral latency because FIV production continues in infected cells in tissues, and virus can still be recovered from blood lymphocytes, serum or plasma, cerebrospinal fluid (CSF), semen, and lymphoid tissues.36,71,357,532 Plasma levels of virus and viral RNA can increase again during the terminal phases of infection, and virus loads may vary from time to time (see Fig. 12-1).236,357,357 FIV infects a wide range of cell types in vitro and in vivo (Table 12-1), but cell tropism is isolate dependent.202,245 Cell tropism is defined by expression of surface proteins that serve as FIV receptors. Unlike HIV, FIV does not use the feline CD4 molecule as a receptor to infect target cells.436,733 Two proteins with receptor function for FIV have been characterized: CD134 and a chemokine receptor designated CXCR4.* CD134 appears to be expressed more in CD4+ cells than in CD8+ cells, B cells, macrophages, or dendritic cells,849 and its level of expression in CD4+ cells is increased with activation,217,849 likely accounting, at least in part, for the well-documented increase in virus production from activated CD4+ cells. Expression of CXCR4 mRNA or antigen can be demonstrated in a large number of cell types known to be susceptible to FIV infection, including lymphocytes, monocytes, macrophages, dendritic cells,849,1024 and astrocytes. CXCR4 has been detected in intraepithelial mononuclear cells and epithelial cells of the rectum, colon, and female reproductive tract, perhaps accounting for some degree of transmucosal or vertical transmission observed in experimental studies.† CD134 may also play a role in cell infection in nondomestic cat isolates.952 The hallmark of FIV pathogenesis is progressive disruption of normal immune function.. Early and persistent immunologic abnormalities that occur after experimental2,46,46 and natural407,737 infection in both domestic and nondomestic felids are decreases in both the number and relative proportions of CD4+ cells (see Fig. 12-1) in the peripheral blood, and in most primary lymphoid tissues examined.126 Causes of CD4+ cell loss may include decreased production secondary to bone marrow or thymic infection, lysis of infected cells induced by FIV itself (cytopathic effects), destruction of virally infected cells by the immune system, or death by apoptosis.* Apoptosis is a form of cell death that follows receipt of a membrane signal initiating a series of programmed intracellular events that ultimately lead to cell death. Apoptosis of CD4+, CD8+, and B cells has been documented in lymph nodes, spleen, and thymus of FIV-infected cats421,899,899; the degree of apoptosis has correlated inversely with CD4+ numbers and the CD4/CD8 ratio.421 Increased expression of proteins that mediate apoptosis, as has been demonstrated in lymphocytes from FIV-infected cats, suggests a mechanism by which lymphocytes are susceptible to apoptotic signals in vivo.127,1036 Proteins coded by the FIV env gene are capable of inducing apoptosis in peripheral blood mononuclear cells by a mechanism that requires CXCR4 binding and may be important for inducing apoptosis in bystander cells.329 Ultimately, loss of CD4+ cells impairs immune responses because CD4+ cells have critical roles in promoting and maintaining both humoral and cell-mediated immunity (CMI). Apoptosis may also contribute to a loss of CD8+ cells on their recognition of FIV antigen.765 New insights into FIV-induced immune dysfunction have emerged with examination of a subset of CD4+ cells termed Treg (for T-regulatory) cells. Under normal conditions, Treg cells are capable of both antigen-specific and nonspecific suppression of immune responses. In FIV-infected cats, Treg cells with suppressive activity have been documented during early infection307,680 and in chronically infected cats.307,789 Treg cells have been shown to inhibit production of interferon (IFN)-γ from CD8+ cells, suggesting that Treg cells could impair immune responses to FIV. Increased activity of Treg cells could also play a role in suppressing immune responses to foreign antigens or pathogens in FIV-infected cats, perhaps accounting in part for some of the susceptibility to infection with other pathogens. In addition, Treg cells are themselves targets for FIV infection,492,493,493 and an inactive form of Treg cells has been shown to support latent FIV infection in vivo, with activation of viral production provoked by mitogen stimulation.494 Thus, Treg cells may serve as a FIV reservoir capable of increasing virus production with appropriate activation or stimulation.494 Experimental inhibition of Treg cells, by use of monoclonal antibody against their CD25hi surface marker allowed FIV-infected cats to produce a more robust humoral response to foreign antigen as compared to cats without Treg cell inhibition.680a Similar to HIV infection in humans, loss of CD4+ cells, in FIV-infected cats, leads to inversion of the CD4/CD8 ratio (see Fig. 12-1), which may occurs weeks to months after infection, depending on the viral isolate studied.2,1018 An increase in the proportion of CD8+ cells, in particular a population referred to as “CD8+ α-hi, β-low cells,” may be contributing to the inverted ratio.* Although inversion of the CD4/CD8 ratio is a consistent feature of both natural and experimental infections, its use as a prognostic tool for cats is questionable, in contrast to its prognostic value in HIV-infected humans.88,1061 FIV-infected cats may show severe inversion for prolonged periods without developing clinical signs, and there is no correlation of the ratio with clinical stage of infection or plasma viremia.358,407 Several other immunologic abnormalities occur with FIV infection.1019 Over time, lymphocytes from infected cats lose the ability to proliferate in response to stimulation with lymphocyte mitogens, or recall antigens, and have impaired priming of lymphocytes by immunogens in vitro.† Lymphocyte function may also be impaired by reduced or altered expression of cell surface molecules such as CD3, CD4, major histocompatibility complex II antigens, other co-stimulatory and signaling molecules, and cytokine receptors.‡ Many of these molecules have a critical role in antigen presentation, or amplification and control of immune responses. FIV infection is associated with disrupted production of cytokines, molecules essential to normal immune function.1036 Cytokine patterns detected from cultures of FIV-infected lymphocytes are dependent to some extent on the FIV isolate and tissue compartment studied.205,611 Reported changes in cytokine production in FIV-infected cats, as compared with noninfected cats, have included increased production of IFN-γ, tumor necrosis factor α, IL-4, IL-6, IL-10, and IL-12.575,599,758,872 Compared to uninfected cats, differences in IL-10/IL-12 ratios have been observed in FIV-infected cats co-infected with Toxoplasma gondii,593 a pattern that could impair development of CMI response to T. gondii. Increased IL-10 also has been documented in cats co-infected with FIV and Listeria monocytogenes, with FIV-infected cats exhibiting delayed clearance and more severe clinical signs of infection as compared with FIV-negative cats.201 Altered IL-10/IL-12 ratios have also been noted in bone marrow–derived dendritic cells in response to stimulation of various toll-like receptors,571 which recognize microbial pathogens and are responsible for initiation of immune responses. Aberrant toll-like receptor responses could also increase the risk for some opportunistic infections. Alterations in function of nonspecific defenses, such as impaired neutrophil adhesion and emigration in response to bacterial products, have been described in FIV-infected cats,372,401,401 and these defects in neutrophil function improved with granulocyte-macrophage colony-stimulating factor (GM-CSF) treatment.401 Natural killer cell activity has been reported as diminished1119 or increased1127 in FIV-infected cats, depending on whether the cats were acutely or asymptomatically infected, respectively. Another manifestation of the immunologic dysregulation observed in many FIV-infected cats is hypergammaglobulinemia, primarily from increases in IgG.2,299 Hypergammaglobulinemia is usually a polyclonal distribution and the IgG produced is not entirely FIV specific. However, it is a direct consequence of FIV infection because clinically healthy FIV-infected, specific-pathogen free (SPF) cats are also hypergammaglobulinemic.299 In addition to increased IgG, increased circulating immune complexes have been detected in FIV-infected cats.660,823 FIV has also been incriminated as causing a delay in the class shift of antibody isotypes from IgM to IgG based on work in cats infected with both FIV and T. gondii.560 Abnormal neurologic function has been described in FIV-infected cats, and the cat remains a model for the study of acquired immunodeficiency syndrome (AIDS)-related neurologic disease in humans. It is clear from all studies that the origin of neurologic signs in FIV-infected cats is multifactorial.* Although results of some studies have linked neurologic dysfunction and neuronal injury with the amount of virus present in the brain of infected cats,483,640 results of many others support the hypothesis that neurologic dysfunction and histologic changes in the central nervous system (CNS) are indirect events and not necessarily an immediate consequence of CNS-cell infection or viral replication within the CNS.672,774,825,886 CNS inflammation is provoked by virus production and increased microglial major histocompatibility complex I expression,398 and traffic of mononuclear inflammatory cells into the brain may be governed by properties of microglial cells and astrocytes.443 CD8+ cell-mediated neurologic injury also has been demonstrated.1128 Experimental infections in vivo have caused brain lesions in the absence of massive CNS infection.886 Neurotoxins such as glutamate have been implicated as one cause of neuronal loss.93,102,835,942,1118 Results of other studies have incriminated a role for matrix metalloproteinases, which can break down collagen and alter properties of the blood-brain barrier, leading to altered neuronal cytoskeletal properties or imbalanced neurotransmitters164,640 in the pathogenesis of neurologic dysfunction in FIV-infected cats.469,480,480 In vitro and, occasionally, in vivo studies suggest that infected brain astrocytes and macrophages may impair normal CNS-cell metabolism, or may be a source of neurotoxins such as inflammatory cytokines or cell-signaling molecules.† Documented abnormalities of astrocyte and macrophage functions include altered intercellular communication, abnormal glutathione reductase activity that may render cells more susceptible to oxidative injury, alterations in mitochondrial membrane potential that disrupt energy-producing capacities of the cell, and impaired maintenance of intracellular calcium concentrations.99 Some studies suggest that sequences in the viral envelope protein and levels of chemokine expression within the CNS are important in neurovirulence properties,‡ and others indicate that neurologic-cell dysfunction does not necessarily correlate with envelope-mediated replicative capacity.82,480 Peripheral nerve dysfunction associated with axonal injury and macrophage activation has also been noted in cats infected with neurovirulent clones of FIV.516 The pathogenesis of some clinical features of FIV remains unexplained but, similar to neurologic disease, may result from abnormal function of, or inflammation in, affected organs. Wasting disease has been observed in the absence of obvious causes such as diarrhea or neoplasia. Abnormal renal function and nephritis have also been reported in FIV-infected cats.586 Many aspects of clinical FIV will reflect the pathogenesis of the secondary diseases such as infections and neoplasms to which FIV-infected cats are considered more susceptible. Experimental FIV infection progresses through several stages, similar to HIV-1 infection in humans. Recognized clinical stages in cats include an acute phase, a clinically asymptomatic phase of variable duration, and a terminal phase of infection often referred to as feline AIDS.274,358 Some investigators have described two other phases in experimentally infected cats in keeping with the terminology for HIV infection: progressive generalized lymphadenomegaly, which follows the asymptomatic phase, followed by AIDS-related complex (ARC).358,465,466,780 Still other researchers describe a sixth category to encompass miscellaneous diseases such as neoplastic, ocular, and immune-mediated diseases that are observed in some infected cats.780 Although division of FIV infection into these clinical stages may prove useful from the standpoint of gauging prognosis, no sharp distinction exists between them, and not all stages will be apparent in most naturally infected cats. Furthermore, there is no means to reliably predict the transition from the asymptomatic phase to the ARC or AIDS phase, although one study found that higher levels of viremia during the acute stage of infection were associated with more rapid progression to the terminal phases of the disease.237 In a study of naturally infected cats, a trend was noted for increasing viremia with progression of clinical signs, with cats in the AIDS category of infection exhibiting higher virus loads than cats in the ARC or asymptomatic phase of infection.358 In contrast to HIV-infected humans, cats that are classified as being in the AIDS phase (high virus load, severe clinical signs due to secondary infection) may recover and may be asymptomatic again and their virus loads may decrease dramatically. Thus, clinical staging, as in HIV infection, is not valuable in FIV-infected cats. Clinical signs of FIV infection are nonspecific. Most clinical signs attributed directly to FIV infection, with the possible exception of FIV-induced neurologic disease, likely go unobserved in many naturally infected cats. During acute experimental infection, clinical signs are usually transient and so mild as to go unnoticed. Some cats exhibit fever and malaise. Signs of acute enteritis, stomatitis, dermatitis, conjunctivitis, and respiratory tract disease have been described in experimentally infected cats. Generalized lymph node enlargement is common during acute experimental infection.739 The acute phase may last several days to a few weeks, after which cats will enter a period in which they appear clinically healthy. The duration of the asymptomatic phase varies and likely depends on factors such as the pathogenic potential of the infecting isolate and the exposure of the infected cats to other pathogens; however, it usually lasts for years. One experimentally infected cat, kept isolated from other cats, had documented viremia for more than 8 years without developing clinical signs.532 The age of the cat at the time of infection may also influence the length of the asymptomatic stage and the severity of clinical disease, depending on the isolate studied.781,812 During the later stages of infection, clinical signs are a reflection of opportunistic infections, neoplasia, myelosuppression, and neurologic disease. Infections with opportunistic pathogens of viral, bacterial (Fig. 12-2), protozoal, and fungal origin have been reported in FIV-infected cats. Few studies, however, have compared the prevalence of most of these infections in corresponding groups of FIV-infected and noninfected cats. In one report, no correlation was found between FIV infection and infection with Cryptococcus or Cryptosporidium.711,1063 Another study reported a higher number of fungal genera isolated from the skin, oropharynx, and rectum of FIV-infected cats compared with noninfected cats, but FIV-infected cats had no signs of fungal infections at the time of examination,647 and no correlation was found between FIV infection and the presence of Cryptococcus neoformans or dermatophytes.941 In one study, antibody prevalence of T. gondii was similar in both FIV-infected and noninfected cats, and FIV-infected cats did not have detectable oocyst shedding.986 In other studies, there has been a high degree of statistical correlation between cats with positive serum FIV-antibody presence and cats with increased T. gondii-specific serum antibody titers.243,627 The prevalence of infection with Bartonella henselae or Bartonella clarridgeiae, the agents of human cat scratch disease, has not been positively associated in cats with FIV infection 124a,350,402,655,656 as it has been in some studies with feline leukemia virus (FeLV) infection.124a In contrast, an increased prevalence of FIV infection has been reported in cats that had serum antibody reactivity to Bornavirus,445 a virus detected in cats in eastern and northern European countries (see Chapter 19), and to orthopoxvirus (see Chapter 17).1028 A statistical association between FIV infection and infection with Mycoplasma haemofelis or Mycoplasma haemominutum60,335,637,991 has also been described, but it is not clear if FIV infection is a true risk factor or if infection with both agents reflects common risk factors (e.g., outside lifestyle, male cats). Results of two studies indicated no effect of preexisting FIV infection on numbers of M. haemofelis, M. haemominutum, or “Candidatus Mycoplasma turicensis” organisms in blood, as determined by PCR.998,1093 Experimentally, FIV infection worsened the respiratory disease observed in an experimental model of acute toxoplasmosis.196 Clinical illness may also be caused by opportunistic infections at sites harboring endogenous microflora or by other secondary pathogens that have not been specified. Respiratory disease may be observed in FIV-infected cats and can result from bacterial, fungal, protozoal, or parasitic infections.57 Diarrhea has been seen in experimentally infected cats in the absence of detectable enteric pathogens. Bacterial overgrowth involving endogenous microflora and inducing inflammatory lesions has been proposed as a possible cause.770 Stomatitis is a common finding in FIV-infected cats and can occur during any stage of infection. The pathogenesis of stomatitis is uncertain, although the histopathologic findings of lymphocytes, plasma cells, and variable degrees of neutrophilic and eosinophilic infiltrates suggest either an immune response to chronic antigenic stimulation or immune dysregulation. Stomatitis has not been a consistent finding with FIV infection (see Chapter 88)846 and is not often seen in experimentally FIV-infected SPF cats, suggesting that exposure to other pathogens may also play a role.586 Co-infection of FIV-infected cats with feline calicivirus, both experimentally and after natural infection, resulted in more severe oral disease (see Clinical Signs, Chapter 14, and Feline Lymphocytic Plasmacytic Ulceroproliferative-Gingivostomatitis (Faucitis), Chapter 88).857,1005 Odontoclastic resorptive lesions have been reported with higher prevalence in cats experimentally infected with FIV, as compared with noninfected cats; such lesions have been speculated to be a consequence of gingivitis or stomatitis present in the cats.408 Neurologic signs have been described in both natural and experimental, and acute and terminal FIV infections.* Neurologic impairment after FIV infection appears to be isolate dependent.834 The most common neurologic signs observed are behavioral changes. Other abnormalities that have been described include seizures, paresis, multifocal motor abnormalities, impaired learning, and disrupted sleep patterns.367,839,839 Neurologic signs may improve if they occur during the acute stage of infection, although residual deficits are possible. Abnormal forebrain electrical activity and abnormal visual and auditory-evoked potentials have also been documented in cats that appeared otherwise normal.52,796,812,814 Less commonly, secondary infections such as feline infectious peritonitis, toxoplasmosis, or cryptococcosis cause the observed neurologic deficits. FIV-infected cats can develop ocular disease,275,559,755,1094 and abnormalities may be found in both anterior and posterior segments. Anterior uveitis may result from secondary infections such as toxoplasmosis or may be directly related to FIV infection.275,755,755 Glaucoma, with and without uveitis, has also been described.275,755,755 Posterior segment changes that may be seen include pars planitis (an infiltration of leukocytes, mainly plasma cells, into the vitreous behind the lens), focal retinal degeneration, and retinal hemorrhages.275,1094 Neoplasia is a common reason that FIV-infected cats are brought to a veterinary clinic. Statistically, FIV-infected cats are much more likely to develop lymphoma or leukemia compared with noninfected cats.144,818 Lymphomas, leukemias, and a variety of other tumor types have been reported in association with FIV infection.* Most lymphomas in FIV-infected cats are B-cell tumors.144,327,818,1006 FIV provirus is only occasionally detected in tumor cells,62–64,1066 suggesting an indirect role for FIV in lymphoma formation, such as decreased cell-mediated immune surveillance or chronic B-cell hyperplasia.64,270 However, clonally integrated FIV DNA was found in lymphoma cells from one cat that had been experimentally infected 6 years earlier,62,63 raising the possibility of an occasional direct oncogenic role of FIV. The prevalence of FIV infection in one cohort of cats with lymphoma was 50%,327 much higher than the FIV prevalence in the population of cats without lymphomas, which is supportive of a cause and effect relationship between FIV and feline lymphoma. In the terminal phase of infection, a wasting syndrome may occur. If experimentally infected with some particularly pathogenic FIV isolates, SPF cats have developed a terminal wasting syndrome within 6 to 8 weeks postinfoculation.235,237
Feline Immunodeficiency Virus Infection
Etiology
Epidemiology
Prevalence
Common Name
Scientific Name
General Topic
Reference
African lion
Panthera leo
Characterization of lion evolutionary history by genetic analysis of lion and FIV
Antunes et al., 200824
Neurologic disease in FIV-positive lions
Brennan et al., 2006104
Serosurvey of viral infection in Ugandan lion
Driciru et al., 2006248
Survey of retrovirus infection in captive lions in Japan
Endo et al., 2004273
Lion strains of FIV use CD134 and CXCR4 as receptors
McEwan et al., 2008669
Examination of FIV genome in lions in Tanzania and Botswana
Pecon-Slattery et al., 2008777
Survey of select infectious disease in lions in Botswana
Ramsauer et al., 2007847
FIV-associated pathology in wild lions
Roelke et al., 2009875
Patterns of variation in several FIV genes and infections with multiple strains among Serengeti lions
Troyer et al., 20041025
Pallas’s cat
Otocolobus manul
Serosurvey of FIV in 28 wild Mongolian Pallas’ cats
Brown et al., 2010113
Puma/cougar
Puma concolor
Population demographics revealed by study of lentivirus
Biek et al., 200675
Assessment of viral load in naturally infected cougars
Blake et al., 200691
Seroprevalence in Rocky Mountain cougars
Biek et al., 200677
FIV recombination in naturally infected cougars
Bruen and Poss, 2007122
Review of molecular biology and evolution of cougar lentivirus
Poss et al., 2008831
Changes in FIV in naturally infected cougars
Poss and Ross, 2008830
Comments on cross-species FIV transmission
Troyer et al., 20081027
Multiple exotic felids
Serosurvey of free-ranging cats in Brazil
Filoni et al., 2006289
Review of evolution of FIV in cats, including several exotic felids
Pecon-Slattery et al., 2008778
Seroprevalence of FIV in 35 felids and 3 hyenids
Troyer et al., 20051026
Bobcat
Lynx rufus
Evaluation of potential transmission of FIV from bobcats to cougars in southern California populations
Franklin et al., 2007315
Reliability of FIV tests for diagnosis of infection in bobcats, ocelots and pumas
Franklin et al., 2007316
Ocelot
Leopardus pardalis
Serosurvey of ocelots on Barro Colorado Island, Panama
Franklin et al., 2008314
Florida panther
Puma concolor coryi
Epidemiology and diagnostic considerations for lentiviruses in Florida panthers
Miller et al., 2006681
Cheetah
Acinonyx jubatus
Serosurvey of 9 feline viruses, including FIV, in free-ranging Namibian cheetahs
Thalwitzer et al., 20101007
Transmission
Pathogenesis
Clinical Findings
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


