Feline foamy virus (FFV), previously known as “feline syncytium-forming virus,” is a retrovirus in the family Retroviridae, subfamily Spumavirinae.24 Host-specific viruses of this genus have been isolated from some mammalian species. Similar to other retroviruses but after or even before infection, FFV produces a DNA copy, using its RNA genome as a template, along with the enzyme reverse transcriptase. This proviral DNA is integrated into the host cell chromosome, and persistent infection, with or without viral production, can result. Spuma or foamy viruses are often highly cell-association and are often have no, or low, pathogenic potential.4 The prevalence of FFV infection is high in both clinically healthy and diseased cats. Virus has been isolated from primary cultures of tissue and body secretions in up to 90% of a population of cats.15 More typically the prevalence of infection in a cat population varies between 25% and 50%, depending on the age, geographic location, and local environment of the cats.* Fifty percent or more of kittens born to FFV-infected queens are infected at birth,30 and 15% of cultures of fetal cats are positive for the virus, suggesting that it can be transmitted vertically.14 In utero infection probably occurs by the transfer of infected maternal leukocytes across the placenta, but infection may also be transmitted through milk of lactating animals.30 In contrast to most feline infectious diseases, the FFV infection rate in cat colonies may be lower than that in the random cat population. Roaming or outdoor cats have the highest FFV prevalence, suggesting possible transmission through bite wounds (like feline immunodeficiency virus [FIV]), but more likely by intimate social contact (like feline leukemia virus [FeLV]).47 This is also supported by serologic evidence that male and female cats, with or without neutering, have an equal rate of positive seroreactivity.29 Some wild cat species also have a relatively high prevalence rate of sera reactive to FFV; however, extensive surveys are not available.7,33 FFV is a nuisance to virologists and manufacturers of feline vaccines because it becomes apparent in many feline tissues and cell cultures that have been subjected to multiple passages, rather than in primary cell cultures.22 In vitro, FFV is not highly host specific; in addition to cat cells, it infects cells of dogs, chickens, humans, and bats.35 When FFV is produced, it is enveloped and has long glycoprotein spikes (Fig. 15-1, A), and viral particles may accumulate in the host cell cytoplasm within membrane-bound vesicles, giving the cell a foamy appearance. In cell culture, it produces multinucleated syncytia within 1 to 2 weeks of growth in certain rapidly multiplying tissue cultures (Fig. 15-1, B). Cell lysis is a late event because large syncytia are not viable. Syncytium formation in cell culture is not unique to FFV; it has also been associated with a variant of FeLV where an insertion into the FeLV envelope glycoprotein gene is responsible for this cytopathic effect.36 Intranuclear inclusions of FFV particles have never been seen; however, cytoplasmic virus is easily visualized by electron microscopy. FFV can be distinguished from FeLV, which is visualized only as it buds from the cell, whereas FFV often forms recognizable preassembled nucleocapsids within the cytoplasm before budding (Fig. 15-1, A). The suitability of FFV as a vector for feline vaccine antigens has been demonstrated.41 The complete replication-competent genome of FFV proviral DNA has been cloned.49,50 It has been shown to be an efficient vector to express vaccine antigens and heterologous genes.3,11,20,21 As a nonpathogenic retrovirus, it appears to be a suitable candidate for use in cats41 or other host species. FFV has never been unambiguously associated with disease. Many cats have been infected naturally and experimentally and had no clinical illness, a scenario that may reflect a very close virus-host coadaptation.39,42 Virus replication appears to be controlled and regulated by the infected cat through the APOBEC3 proteins and other restriction factors as part of the intrinsic immunity.25,28 Reports about FFV pathogenicity are controversial and conflicting. The presence of the virus in 100% of cats affected with chronic progressive polyarthritis has been reported.32 Concurrent FeLV infection was found in 70% of these cats. The prevalence of infection with both viruses was 2 to 10 times greater than that in age-matched cats not having chronic progressive polyarthritis. By altering the host immune system, FeLV may potentiate the ability of FFV to produce disease. Combined infections with FFV and FIV are also frequently found.2 However, co-infection with FFV and FIV did not increase the severity of FIV-induced illness in the early stages of the disease.50 A combined infection is most likely a result of a common mode of transmission rather than a mutual pathogenic mechanism. In vitro FFV strains infect lymphoblastoid cells, producing altered replication, syncytia formation, and fragmentation of cellular DNA.18 These features of apoptosis may be responsible for independent immune alterations in infected cats. Most FFV-infected cats are asymptomatic. Chronic progressive polyarthritis of cats generally affects naturally infected males between
Feline Foamy (Syncytium-Forming) Virus Infection
Etiology
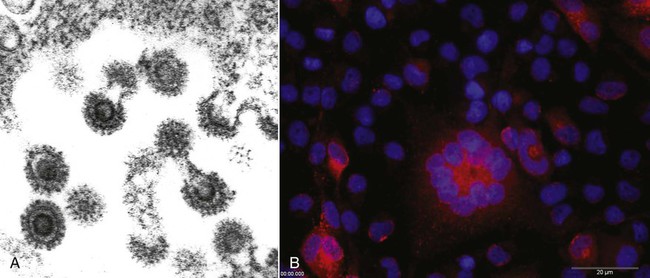
Pathogenesis
Clinical Findings
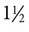 and 5 years of age.27,31,32,44 Two forms of the disease have been described: one with osteoporosis and periarticular periosteal proliferation, the other with periarticular erosions, collapse of the joint space, and joint deformities. Lymphadenomegaly, swollen joints, and stiff gait are caused by both types. In experimentally FFV-infected cats that were monitored for a 6-month period postinfection, histopathologic changes of mild glomerulonephritis and moderate interstitial pneumonia were observed10; however, no corresponding cohoused uninfected control cats were evaluated to substantiate the association of the virus infection with these mild lesions.
and 5 years of age.27,31,32,44 Two forms of the disease have been described: one with osteoporosis and periarticular periosteal proliferation, the other with periarticular erosions, collapse of the joint space, and joint deformities. Lymphadenomegaly, swollen joints, and stiff gait are caused by both types. In experimentally FFV-infected cats that were monitored for a 6-month period postinfection, histopathologic changes of mild glomerulonephritis and moderate interstitial pneumonia were observed10; however, no corresponding cohoused uninfected control cats were evaluated to substantiate the association of the virus infection with these mild lesions.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


