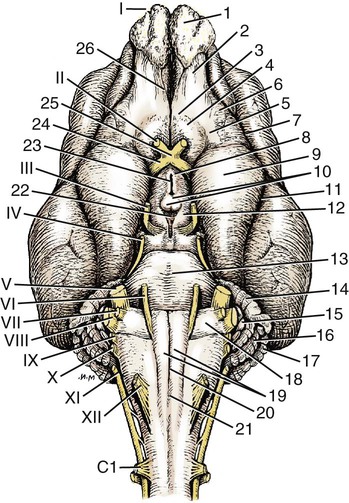
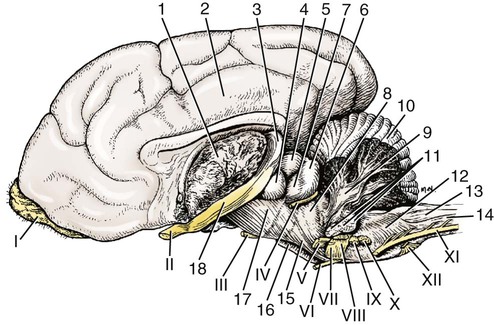
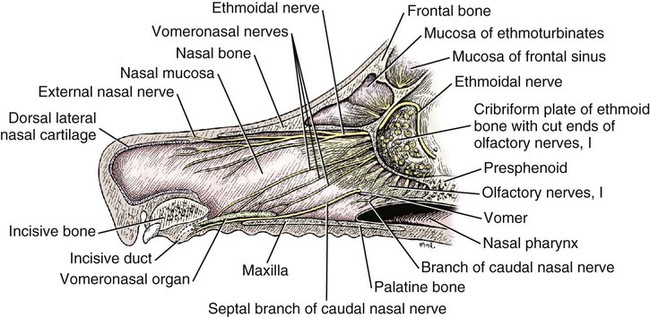
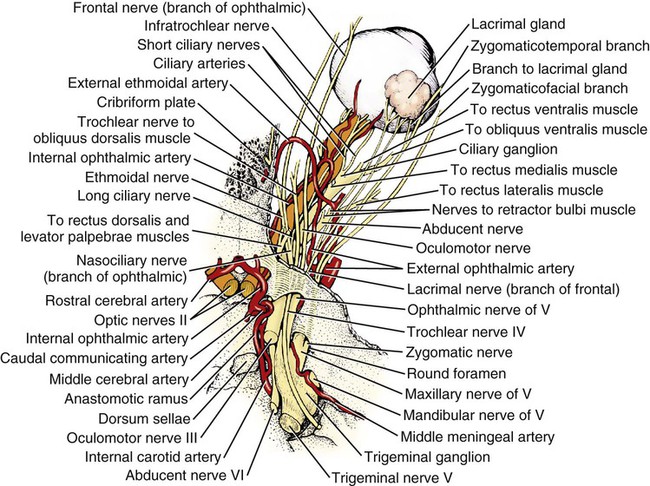
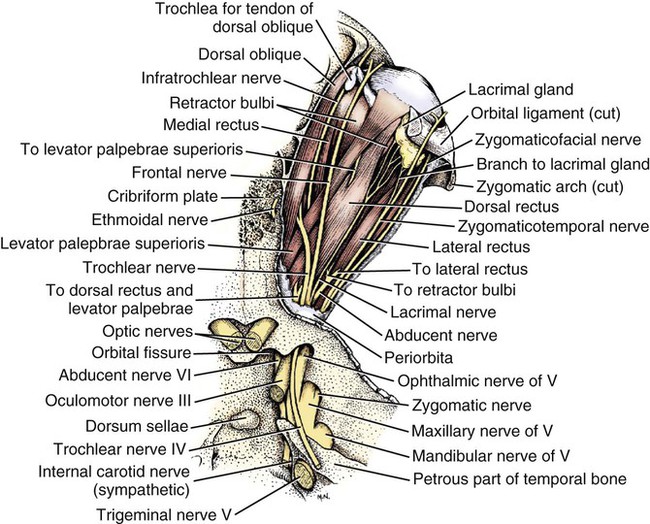
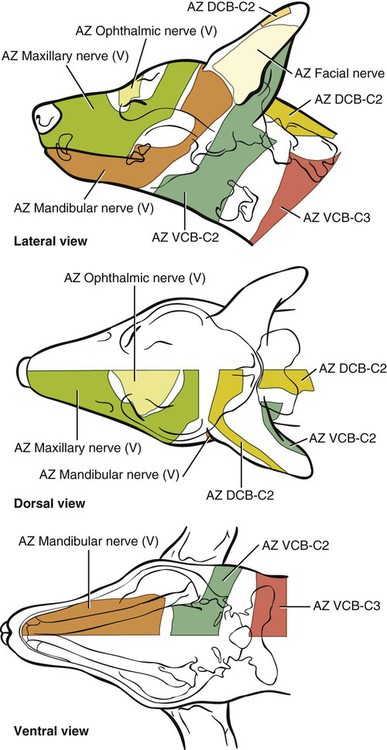
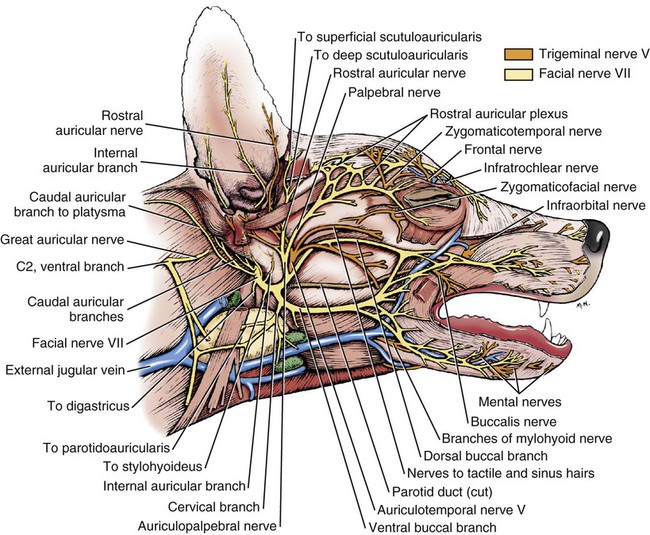
The cranial nerves of vertebrates have been the subject of much study because of their association with the sense organs of the head, their easy accessibility, and their interesting phylogenetic and ontogenetic history (Kappers et al., 1936). From a practical standpoint they have clinical relevance as diagnostic indicators of central nervous system (CNS) disorders (de Lahunta, 1983). It is probable that most of the cranial nerves were once associated with a pattern of head segmentation, not unlike dorsal and ventral segmental spinal nerves. In fish and amphibians there are only 10 pairs of cranial nerves, because the succeeding nerves are not enclosed by the skull and hence are considered as spinal nerves. The first spinal nerve of fish is large and serves gill and opercular structures that are destined to be incorporated into the skull. Not until reptiles evolved did the development of the skull envelop the first and second spinal nerves, making them cranial nerves XI (accessory nerve) and XII (hypoglossal nerve). The cranial nerve innervation of some ventral neck muscles and the trapezius muscle to the scapula can be understood only in the context of their phylogenetic history beginning as the gill arch levator muscles of fish (Romer & Parsons, 1986). The ontogenetic sequences of muscle-nerve relations often reflect their phylogenetic history (Noden & de Lahunta, 1985). Reptiles, birds, and mammals all have 12 pairs of cranial nerves, which by convention are numbered with Roman numerals I to XII. The cranial nerves were named according to early functional interpretations in humans. Long after the present numbering scheme was adopted, an additional, accessory olfactory nerve, the nervus terminalis, was recognized as a vomeronasal component of the olfactory system and was sometimes given the numeral designation of cranial nerve “0.” Locy (1899), when first describing this nerve, suggested a neural crest and placodal origin. Contrary to statements that sometimes appear in the literature as to the absence of the terminal nerve in humans, it is present and has been described by Johnston (1914), Brookover (1914), and others. It is considered here as a part of cranial nerve I, the olfactory nerve. For other overall considerations of the cranial nerves and adnexa of the dog, the reader is referred to Jenkins (1972), McClure (1979), Meyer (1979), and de Lahunta and Glass (2009). The cranial nerves are listed in Box 19-1. The olfactory nerves (nn. olfactorii), usually referred to collectively as the first cranial nerve, consist of numerous nonmyelinated axons with cell bodies located in the olfactory epithelium covering one half of the ethmoidal labyrinth and the dorsal part of the nasal septum. Axons from these olfactory cells enter the skull through the cribriform plate of the ethmoid (Fig. 19-1), deep in the caudal part of the nasal cavity, to reach the olfactory bulbs of the brain where they synapse. The axons do not form a discrete nerve trunk, because they enter the bulbs through many cribriform foramina over the entire contact surface (ventral surface) of the olfactory bulbs. The olfactory nerves are usually torn from the brain when it is removed from the skull. Included with the olfactory nerves are the terminal nerve (n. terminalis) and the vomeronasal nerve (n. vomeronasalis) both of which arise from the vomeronasal organ (Fig. 19-1) and which enter the olfactory peduncle and bulb, respectively, and form a distinct bundle and tract (Scalia & Winans, 1976). Read (1908) studied the gross and histologic structure of the olfactory mucosa, vomeronasal organ, and olfactory nerves in the dog, cat, and human. Her historical review points out some of the past controversy involving this relatively simple sensory complex and is followed by an investigation with the techniques of that period. She used 20% nitric acid for 6 to 12 hours to decalcify the bone so that it could be removed from the mucosa to expose the nerves. The nerves appeared white on a dark background of mucosa. Dissection was done under water in bright light, which allowed very small nerves to be seen and photographed. This gross technique is still applicable today but is little used. The epithelium of the olfactory region shows three kinds of cells: sustentacular cells, which are elongate and cylindrical; olfactory cells, which are fusiform and lie between sustentacular cells; and stellate or basal cells near the basement membrane. In the submucosa there are serous glands. Fresh olfactory mucosa is slightly yellowish in appearance owing to pigment in the sustentacular cells. The olfactory nerve cell bodies are large and numerous (Fig. 19-1), and all of the folds of mucosa in the region of the cribriform plate are olfactory. Read (1908) concluded that the vomeronasal organ was intimately connected with the sense of smell, and we now know that it serves as a pheromone receptor for odors associated with estrus and reproductive functions. Also seen within the olfactory mucosa is the ethmoidal nerve (n. ethmoidalis), a branch of the ophthalmic nerve (V), which enters from the orbit through the rostral ethmoidal foramen into the cranial cavity and thence through the cribriform plate into the nasal cavity (Figs. 19-1 and 19-2). The ethmoidal nerve passes along the dorsal border of the nasal septum before dividing into three branches. The external nasal branches (rami nasales externi) pass to the skin in the vestibule of the nose (Whalen & Kitchell, 1983a). The medial nasal branch (ramus nasalis medialis) innervates the septum and the lateral nasal branch (ramus nasalis lateralis) innervates the nasal conchae. Stimulation of the receptors of this nerve, as well as of the nasal branches of the maxillary (V) nerve, lead to sneezing. The caudal nasal nerve (n. nasalis caudalis) from the maxillary nerve (V) enters the nasal cavity through the sphenopalatine foramen and innervates the nasal mucosa and maxillary recess rostral to the ethmoidal labyrinth. The nasopalatine nerve (n. nasopalatinus) from the maxillary (V) is a continuation of the caudal nasal nerve that passes to the incisive canal and vomeronasal organ. McCotter (1913) described the terminal nerve (n. terminalis) in the dog and cat and considered the relationship of the vomeronasal nerves to the olfactory nerves in several mammals (McCotter, 1912). Larsell (1920) reviewed the history of the discovery of the terminal nerve and also described its structure in several mammals, including the dog. The nervus terminalis passes through the cribriform plate in company with vomeronasal bundles. Only one small ganglion was found in the dog by Larsell (1920). It was fusiform in shape and located within the cranial cavity on the ventrolateral surface of the olfactory bulb. Barone et al. (1966) in their review of Jacobson’s organ noted the difficulty they had observing the terminal nerve. They cite 59 papers relating to the vomeronasal organ and its nerves, several of which refer to the dog. The optic nerve (n. opticus), or second cranial nerve, is actually a tract of the brain and not a nerve by definition, but it is called a nerve by convention. Its developmental origin from ganglion cell axons in the optic vesicle, its neuroglial cells, lack of Schwann cells, myelin produced from oligodendrocytes and its meninges with a subarachnoid space are evidence of its CNS status. The optic nerve axons collect at the optic disc of the retina, where they are myelinated, pass through the cribriform area of the sclera, and enter the skull through the optic canals of the presphenoid bone. In all mammals a majority of the axons cross to the opposite side (decussate) at the optic chiasm rostral to the hypophysis (Fig. 19-3) before forming the optic tracts on the lateral aspects of the diencephalon (Fig. 19-4). The axons that cross originate largely from the medial or nasal aspect of each retina. Approximately 75% of the fibers cross in the dog. 1. Cut surface between cerebrum and brainstem (internal capsule) 2. Medial surface of right cerebral hemisphere 7. Brachium of caudal colliculus 9. Corpus medullare of cerebellum 12. Superficial arcuate fibers 13. Spinal tract of trigeminal nerve 14. Dorsal spinocerebellar tract 15. Longitudinal fibers of pons The chiasma, or cross-over, is continued by the optic tract (Fig. 19-4), which arches around the diencephalon and terminates in the lateral geniculate nucleus, pretectal region, and rostral colliculus. The oculomotor nerve (n. oculomotorius), or third cranial nerve, consists primarily of general somatic efferent neurons that innervate several of the striated, voluntary, extraocular muscles that have developed from head somitomeres and parasympathetic general visceral efferent neurons that innervate ocular smooth muscle. The cell bodies of the general somatic efferent neurons are located in the oculomotor nuclei of the rostral mesencephalon, which lie adjacent to the midline on the ventral border of the central grey substance ventral to the mesencephalic aqueduct. The cell bodies of parasympathetic preganglionic general visceral efferent neurons are located in the parasympathetic nucleus of the oculomotor nerve, which lies rostral to the somatic efferent nucleus. As the axons of the oculomotor nerve pass ventrally through the reticular formation of the tegmentum, some fibers cross to the opposite side but most remain uncrossed. The actual proportions are not known for the dog. In the monkey, according to Warwick (1953), the medial and ventral rectus and the ventral oblique muscles are supplied by uncrossed fibers only; the dorsal rectus receives only crossed fibers, whereas the levator palpebrae superioris muscle has a bilateral innervation. The oculomotor nerves emerge from the mesencephalon in the interpeduncular fossa (Fig. 19-3) medial to the crus cerebri. Each nerve courses rostrally through the middle cranial fossa, lateral to the hypophysis and dorsal to the cavernous sinus and passes through the orbital fissure (Fig. 19-5). On entering the orbit, the oculomotor nerve divides into a small dorsal branch and a large ventral branch. The dorsal branch supplies the dorsal rectus and then penetrates that muscle to go dorsally to supply the levator palpebrae superioris muscle. The ventral branch continues rostrally, lateral and slightly ventral, to the optic nerve, where it terminates in a number of branches to the medial rectus, ventral rectus, and ventral oblique muscles. At its termination is the small ciliary ganglion (ganglion ciliare) (Figs. 19-5 to 19-7) where the preganglionic parasympathetic fibers synapse with neuronal cell bodies of postganglionic axons (Fig. 19-7). These postganglionic axons compose the short ciliary nerves that course along the optic nerve to enter the eye and innervate the ciliary muscle (which affects the curvature of the lens) and the sphincter muscle of the iris, m. dilator pupillae, which regulates the amount of light passing through the pupil. The trochlear nerve (n. trochlearis), or fourth cranial nerve, provides general somatic efferent innervation to the dorsal oblique muscle of the contralateral side from its cell bodies of origin. It is unique in two ways: It is the only cranial nerve that emerges from the brainstem dorsally (Fig. 19-4), and it is the only cranial nerve to cross entirely to innervate a muscle on the contralateral side. Its cell bodies lie in the trochlear nucleus of the caudal mesencephalon at the level of the caudal colliculi adjacent to the midline on the ventral border of the central gray substance surrounding the mesencephalic aqueduct. The trochlear nucleus is caudal to the oculomotor nucleus in the caudal part of the mesencephalon. The axons of the trochlear nerve course dorsally around the mesencephalic aqueduct, enter the thin rostral medullary velum caudal to the caudal colliculi, and cross at the trochlear decussation (decussatio nervorum trochlearium) in the velum to the opposite side. The nerve emerges from the velum caudal to the contralateral colliculus, passes rostroventrally over the side of the mesencephalon, and leaves the cranial cavity through the orbital fissure to enter the periorbita and innervate the dorsal oblique muscle of the eyeball (Figs. 19-5, 19-6, and 19-8). The trigeminal nerve (n. trigeminus), or fifth cranial nerve, has both motor and sensory components. The sensory portion is larger. The nerve penetrates the pons (Figs. 19-3 and 19-4) just caudal and ventral to where the transverse fibers of the pons are continued dorsally as the middle cerebellar peduncle. This is rostral to the lateral end of the trapezoid body. The trigeminal nerve enters the trigeminal canal (Fig. 19-8) on the rostromedial aspect of the petrosal part of the temporal bone. Within this canal is the large trigeminal ganglion (ganglion trigeminale) (Figs. 19-5 and 19-9) that contains sensory cell bodies of the general somatic afferent axons found in all three branches of this nerve. As the trigeminal nerve emerges from the trigeminal canal it divides into three nerves: (1) ophthalmic (sensory), (2) maxillary (sensory), and (3) mandibular (sensory and motor). The major branches of these three nerves are shown in Figure 19-10. Using electrophysiologic techniques, Whalen and Kitchell (1983a, b) determined the cutaneous areas of the head and face innervated by all cutaneous nerves of the head. The cutaneous areas of the three major nerves from the trigeminal nerve, including the autonomous zones (the area innervated by only one nerve) and the overlap zones, are shown in Figure 19-11. The ophthalmic nerve (n. ophthalmicus) consists of general somatic afferent axons from the eyelids, eyeball, nasal mucosa, and skin of the nose. It passes through the orbital fissure of the skull (Fig. 19-8). Within the periorbita, the ophthalmic nerve divides into three branches—the frontal nerve, lacrimal nerve, and nasociliary nerve. The frontal nerve (n. frontalis) is the most dorsal of the three branches of the ophthalmic nerve. It passes rostrally (Figs. 19-5 and 19-8) in the periorbita dorsal to the dorsal oblique and dorsal rectus muscles of the eyeball. The frontal nerve becomes subcutaneous just caudal to the orbital ligament to terminate by dividing into supraorbital (n. supraorbitalis) and supratrochlear (n. supratrochlearis) nerves (Figs. 19-10 and 19-12). These nerves collectively supply a cutaneous area on the lateral two thirds of the superior eyelid continuing to the dorsal midline (Fig. 19-13). The cutaneous area overlaps rostrally with the infratrochlear branch of the nasociliary nerve (ophthalmic V) and caudally with branches of the maxillary and mandibular nerves. The lacrimal nerve (n. lacrimalis) is smaller than the other two branches and runs along the lateral rectus muscle of the eyeball (Figs. 19-6 and 19-8) to supply the lacrimal gland. It carries postganglionic parasympathetic axons that have their cell bodies in the pterygopalatine ganglion (see discussion of the facial nerve and Figure 19-7). The nasociliary nerve (n. nasociliaris) is the largest and most medial branch of the ophthalmic nerve. It runs medially deep in the orbit across the dorsal surface of the retractor bulbi muscle (Fig. 19-6). The nerve sends a communicating branch to the ciliary ganglion (ramus communicans cum ganglio ciliari) (Fig. 19-7). This branch contains postganglionic sympathetic axons that bypass the ciliary ganglion to enter the short ciliary branches of the oculomotor nerve (Fig. 19-5). These postganglionic axons have their cell bodies located in the cranial cervical ganglion. They reach the ophthalmic nerve by means of an internal carotid plexus around the internal carotid artery (Fig. 19-20), which go with the artery into the carotid canal via the tympanooccipital fissure, then enter the middle cranial fossa of the cranial cavity via the foramen lacerum. Here they join the ophthalmic nerve ventrally as it branches from the trigeminal ganglion. These axons supply the smooth orbitalis muscle in the periorbita and the three eyelids and the dilator of the pupil. The orbitalis muscle tonically retracts the superior, inferior and third eyelids and protrudes the eyeball. The nasociliary nerve then gives off a number of long ciliary nerves (nn. ciliares longi) that enter the eyeball dorsal and medial to the optic nerve (Fig. 19-6). These branches are the only general somatic afferent branches to the eyeball, including the corneal epithelium and the bulbar conjunctivum. The nasociliary nerve then terminates by dividing into ethmoidal and infratrochlear branches. The ethmoidal nerve (n. ethmoidalis) turns medially (Figs. 19-5 and 19-6) to pass through the rostral ethmoidal foramina and the cribriform plate (Fig. 19-1) to enter the nasal cavity, where it divides into medial, lateral, and external branches. The medial nasal branch (ramus nasalis medialis) supplies the septum and parts of the wall of the nasal cavity (Fig. 19-1). The lateral nasal branch (ramus nasalis lateralis) innervates the mucosa of the nasal conchae. The external nasal branches (rami nasales externi) (Fig. 19-1) innervate the skin of the vestibule of the nostril (Whalen & Kitchell, 1983a) (Fig. 19-2). The infratrochlear nerve (n. infratrochlearis) runs along the medial aspect of the orbit to emerge subcutaneously just ventral to the trochlea of the tendon of the dorsal oblique muscle near the medial canthus of the eye (Fig. 19-12). It supplies an area surrounding the medial canthus (Fig. 19-13). Its cutaneous area overlaps with the frontal nerve dorsally and with branches of the maxillary nerve ventrally. The maxillary nerve (n. maxillaris) is sensory to the superior eyelid, nasal mucosa, superior teeth, superior lip, and nose (Fig. 19-14), and its distal branches contain postganglionic parasympathetic axons that innervate the lacrimal, nasal, and palatine glands (Fig. 19-7). It leaves the cranial cavity through the round foramen, alar canal, and rostral alar foramen (Figs. 19-5, 19-8, 19-9) and courses rostrally on the dorsal surface of the medial pterygoid muscle to the maxillary foramen (Figs. 19-15 and 19-16). In the orbit on the dorsal surface of the medial pterygoid muscle ventral to the periorbita, the maxillary nerve gives rise to the zygomatic nerve, pterygopalatine nerve, and infraorbital nerve (Fig. 19-10). The zygomatic nerve (n. zygomaticus) arises from the maxillary nerve just proximal to the point of entry of the maxillary nerve into the round foramen (Fig. 19-5), or within the alar canal. After the zygomatic nerve exits from the rostral alar foramen (Fig. 19-16), it penetrates the periorbita and divides into zygomaticofacial (ramus zygomaticofacialis) and zygomaticotemporal (ramus zygomaticotemporalis) branches (Figs. 19-5, 19-8, and 19-16). The zygomaticotemporal branch is the more dorsal branch, which runs rostrally in the orbit deep to the periorbita to emerge subcutaneously lateral to the orbital ligament (Fig. 19-8), where it curls ventrally and laterally around the zygomatic arch (Fig. 19-10) to ramify in the skin (Fig. 19-12). It supplies a cutaneous area that includes all of the inferior eyelid (Fig. 19-14). It is overlapped at the lateral canthus of the eye by the frontal nerve (ophthalmic V) (Fig. 19-13) and at the medial canthus by the infratrochlear nerve (ophthalmic V) (Fig. 19-13). The zygomaticofacial nerve is the more ventral branch, which runs rostrally in the orbit deep to the periorbita to emerge medial to the orbital ligament (Fig. 19-8), where it curls dorsally (Fig. 19-12) on the external surface of the temporalis muscle. It supplies a cutaneous area dorsal to the zygomatic arch, over the external surface of the temporalis muscle cranial to the external ear (Fig. 19-14). It overlaps the cutaneous area of the frontal nerve, dorsal to the lateral canthus of the eye. The lateral canthus of the eye is in the overlap zone of the frontal (ophthalmic V) and of the zygomaticofacial (maxillary V) nerves (Figs. 19-13 and 19-14). The pterygopalatine nerve (n. pterygopalatinus) arises from the deep surface of the maxillary nerve on the dorsal surface of the medial pterygoid muscle (Figs. 19-15 and 19-16). The pterygopalatine ganglion (discussed with the facial nerve) lies dorsal to the pterygopalatine nerve and gives off postganglionic parasympathetic axons that join the lacrimal branch of the ophthalmic (V) and zygomaticotemporal nerve (maxillary V) to supply the lacrimal gland (Figs. 19-7 and 19-8). Other branches from the pterygopalatine ganglion go to the following nerves to convey postganglionic parasympathetic axons to the nasal and palatine glands. The minor palatine nerve (n. palatinus minor) arises from the pterygopalatine nerve and curls around the rostral border of the medial pterygoid muscle (Figs. 19-15 and 19-16) to reach the soft palate. In addition to supplying general visceral afferent axons to the soft palate, this nerve contains the postganglionic parasympathetic axons referred to previously and special visceral afferent axons for taste that have their cell bodies in the geniculate ganglion of the facial nerve (discussed with the facial nerve). These taste buds are in the mucosa of the ventral surface of the soft palate. Their special visceral afferent axons travel to the geniculate ganglion through the pterygopalatine ganglion and the major petrosal nerve. The major palatine nerve (n. palatinus major) arises from the pterygopalatine nerve distal to the minor palatine nerve (Figs. 19-5 and 19-16). The major palatine nerve gives off an accessory palatine nerve (n. palatinus accessorius), which supplies the caudal part of the hard palate (Fig. 19-2). The major palatine nerve provides a branch to the caudal nasal nerve and continues rostrally to enter the palatine canal via the caudal palatine foramen. It emerges on the hard palate from the major palatine foramen and supplies most of the mucosa of the hard palate (Fig. 19-2). The caudal nasal nerve (n. nasalis caudalis) is the continuation of the pterygopalatine nerve. It lies dorsal to the major palatine nerve in the pterygopalatine fossa (Figs. 19-2, 19-15, and 19-16). The caudal nasal nerve leaves the fossa via the sphenopalatine foramen to enter the nasal cavity (Figs. 19-1 and 19-2). It gives off a small branch to supply the maxillary recess (Fig. 19-2), then the major part of the nerve continues to run rostrally to give sensory innervation to the nasal mucosa surrounding the ventral meatus of the nasal cavity. In addition, it contains postganglionic parasympathetic axons from cell bodies located in the pterygopalatine ganglion (Fig. 19-7). These axons supply the nasal glands. The infraorbital nerve (n. infraorbitalis) is a direct continuation of the maxillary nerve after the caudal nasal nerve separates from the maxillary (Fig. 19-16). In the pterygopalatine fossa, the infraorbital nerve gives off the caudal superior alveolar branches (rami alveolares superiores caudalis) (Figs. 19-15 and 19-16). These arise from the ventral aspect of the infraorbital nerve and run rostroventrally to enter the alveolar canals via the alveolar foramina of the maxilla to supply the caudal superior cheek teeth. An alveolar canal leads to the tip of each alveolus to enter the root of each tooth. After the caudal superior alveolar branches are given off, the infraorbital nerve enters the infraorbital canal by passing through the maxillary foramen. Once within the canal, the infraorbital nerve gives off from its ventral border, the middle superior alveolar branches (rami alveolares superiores medi), which enter alveolar canals via alveolar foramina to supply the superior cheek teeth. Just before the infraorbital nerve exits from the infraorbital canal through the infraorbital foramen, it gives off from its ventral surface the rostral superior alveolar branches (rami alveolares superiores medii) (Fig. 19-10). These branches enter the incisivomaxillary canals to supply the superior canine and incisor teeth. External to the infraorbital foramen, the infraorbital nerve divides into a number of fasciculi, which are named external nasal branches (rami nasales externi), internal nasal branches (rami nasales interni), or superior labial branches (rami labiales superioris), depending on the area supplied (Figs. 19-1, 19-12). The cutaneous area for the infraorbital nerve is shown in Figure 19-14. It is overlapped by other branches of the maxillary nerve (Fig. 19-14) and by the infratrochlear nerve (ophthalmic V) ventral to the medial canthus of the eye (Fig. 19-13).
Cranial Nerves
Olfactory Nerve (Cranial Nerve I)
Optic Nerve (Cranial Nerve II)
Oculomotor Nerve (Cranial Nerve III)
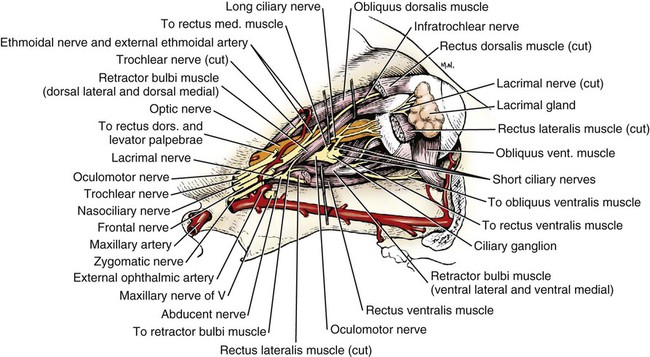
Trochlear Nerve (Cranial Nerve IV)
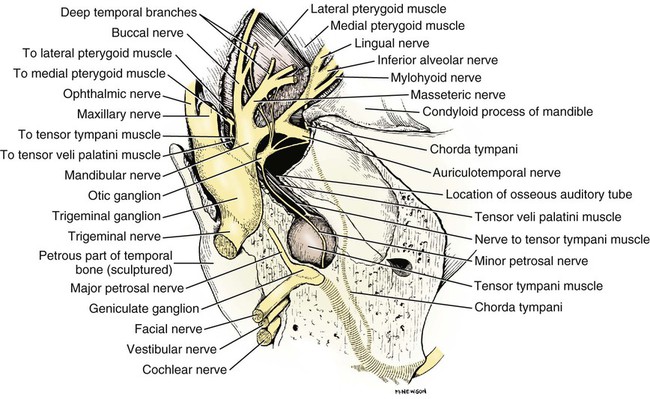
Trigeminal Nerve (Cranial Nerve V)
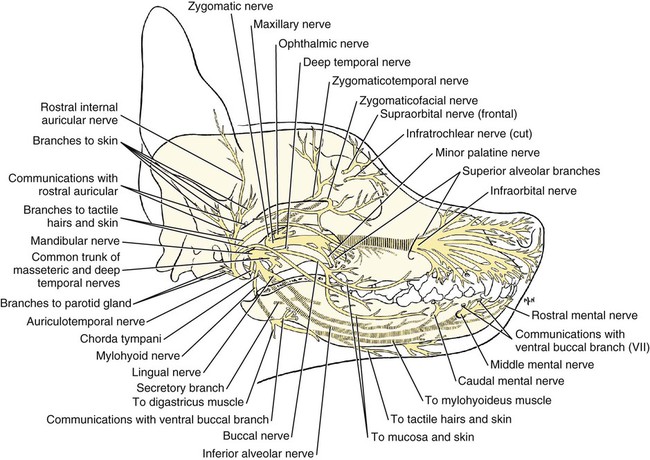
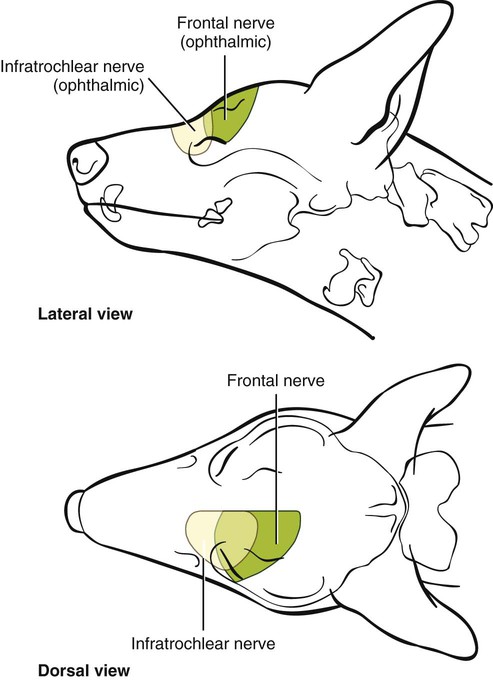
The Ophthalmic Nerve
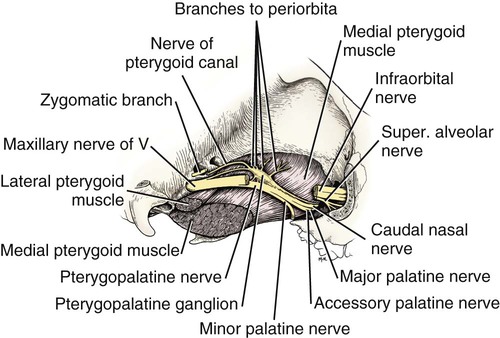
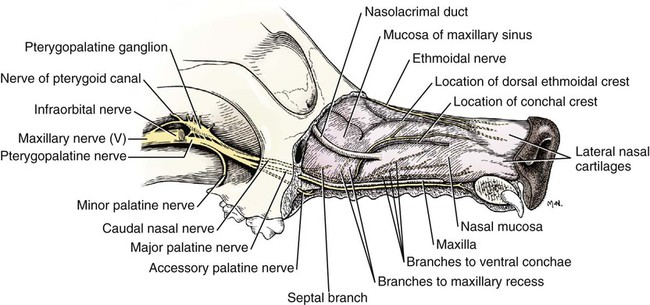
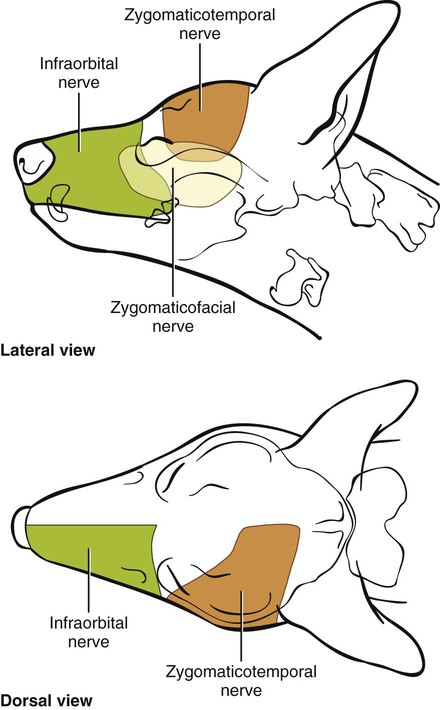
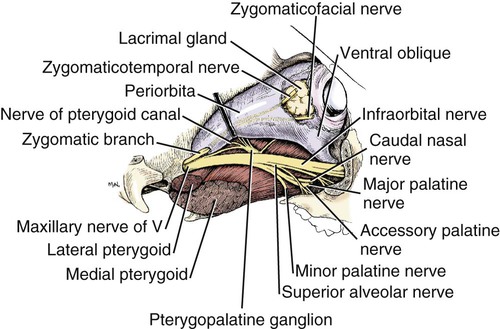
The Maxillary Nerve
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


