Chapter 12 Basic Anatomy, Physiology, and Husbandry
Etymology
The mammalian order Lagomorpha consists of two living families: Leporidae (rabbits and hares) and Ochotonidae (pikas) (Fig. 12-1). The scientific name for the Old World or European rabbit from which all domestic breeds originate is Oryctolagus cuniculus. The genus name is derived from the words orukter (Greek: a tool for digging) and lagos (Greek: a hare) and the species name is derived from cuniculus (Latin: a rabbit or an underground passage). In contrast to most other species in Lagomorpha, rabbits are burrowing animals.
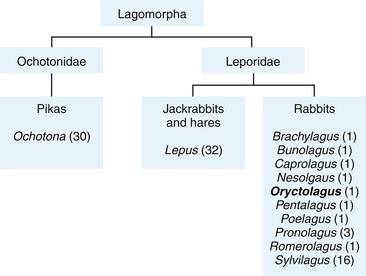
Through human mediation, wild European rabbits have been dispersed to all continents except Antarctica. This has led to some confusion with the common name rabbit. The family Leporidae splits broadly into two groups: the hares of the genus Lepus, containing 32 species, and the rabbits in the remaining 10 genera, which include Oryctolagus (Fig. 12-2). However, these vernacular names are often used synonymously and applied to the wrong animals. For example, the African red rock hares (Pronolagus species) and the endangered hispid hare (Caprolagus hispidus), although commonly called hares, are rabbits. Some hares are called rabbits—for example, the snowshoe rabbit (L. americanus) and, in North America, the several species of hare known as jackrabbits (shortening of jackass-rabbit, so called for its long ears). Even more confusing, in North America, cottontails, or New World rabbits, Syvilagus species (11 species), are commonly called rabbits. Both the European rabbit and the eastern cottontail (S. floridanus) are unique because of their great diversity of habitat: fields, farms, woodlands, deserts, swamps, and forests. However, in North America, the European rabbit Oryctolagus cuniculus has not become feral and is found only as a domesticated animal; all wild rabbits in North America are cottontails.

The English word rabbit (from Middle English rabbet, from Middle French dialect rabbotte, from Walloon robète [baby rabbit], diminutive of Middle Dutch robbe) arose in the fourteenth century to mean the young of the species. The adult was a coney (from Anglo-Norman conis, plural of conil from Latin cunicula) until the eighteenth century and the name can be found in the King James Bible (Leviticus 11:5; Proverbs 30:26). Baby rabbits were an important source of food in the early Middle Ages, because Pope Gregory I (c. 540-604) officially classified laurices (unborn or newly born rabbits) as “fish” and thus permissible to eat during Lent and other Christian fasts. In sixteenth-century British slang, a coney was someone to cheat or rob and doing so was known as “coney-catching”19; the Lancashire expression “coney-fogle” meant “to lay plots.”8 Although the pronunciation of coney (rhyming with honey and money) was changed from a short to long vowel (rhyming with boney),18 the word rabbit gradually pushed out coney in common usage. The similarity of present-day European words for rabbit to the English coney can be seen: coniglio [Italian], conejo [Spanish], konijn [Dutch], Kaninchen [German]. While there are specific words in English to mean the young of a species (e.g., dog/puppy, hare/leveret, cat/kitten) there is no word for the young of rabbits, although they are often referred to as kits or by the hypocoristic term bunny.
Taxonomy and Similarities to Rodents
Most veterinarians observe that rabbits and rodents show a similarity to each other. In these animals, the anterior incisors in the upper and lower jaws are modified to form chisel-like cutting organs.36 Enamel is deposited on the anterior surfaces only, the back surface being dentin. Because enamel is harder than dentin, the front surface wears down more slowly and the incisors remain in a permanently sharp condition from gnawing. This modification is associated with a loss of teeth between the last incisor and the first cheek tooth so that there is a toothless interval referred to as the diastema.
Karl von Linne (also called Linnaeus, 1717-1738), who originated the systematic classification of plants and animals, recognized the likeness of rabbits and rodents to each other. In his Systema Naturae (10th ed., 1758) he assigned rabbits and rodents to a group called Glires (Latin glis = dormouse). Later naturalists designated Glires as the mammalian order Rodentia. Within this order rabbits, hares, and pikas were grouped in the suborder Duplicidentata because they possessed a second pair of incisor teeth in the upper jaw. Most rodents, which have only a single pair of upper incisors, were grouped in the suborder Simplicidentata. However, throughout the twentieth century there has been a strong consensus that the similarities of rabbits and rodents exemplify convergent evolution (i.e., the development of a similar adaptive morphological trait by two or more unrelated species). Consequently naturalists now designate the Duplicidentata as the mammalian order Lagomorpha (to which rabbits belong) and restrict the order Rodentia to the larger group of mammals with only one pair of upper incisors, such as squirrels, rats, mice, and guinea pigs. Based on results of DNA work, in recent years the concept that the two orders are in fact related has resurged. The term Glires, which Linnaeus originally used, is now used to describe the infraclass that encompasses these two orders.
Breeds and Varieties
Rabbits are divided into over 60 fancy breeds and fur breeds. The fur group is divided into normal fur breeds, Rex breeds, and Satin breeds. The normal fur breeds have a coat made up of an undercoat and projecting guard hairs; the Rex breeds have short guard hairs that do not appear above the level of the undercoat; and the Satin breeds have an abnormal hair fiber that produces a sheen.71 The term variety describes a color (e.g., black, blue, steel gray, tortoiseshell) within a breed. Over 500 varieties are described. For a list and images of domestic rabbit breeds, refer to the American Rabbit Breeders Association (www.arba.net) or the British Rabbit Council (www.thebrc.org).
Body conformation and ear size vary widely among breeds of rabbits, and rabbit fanciers have coined some unusual terms to describe lagomorph body shape and fall of ear. They refer to the small, chunky body of a dwarf rabbit, like a cobblestone, as cobby; they describe the long, lean body of a Belgian hare as racy; and they often describe giant rabbits as mandolin-shaped because of the high, curved top line over their hindquarters.81 Most breeds of rabbits have upright ears, which can be long or short. However, some breeds have soft, pliable ears that hang downward and are incapable of erection; we know these as lops.
Life Span
Records for the life span of rabbits in captivity are 9, 10, and 18 years.13 Veterinarians often report seeing pet rabbits 9 to 10 years of age, and one of the authors has seen a 15-year-old pet rabbit. Wild rabbits in an enclosure and receiving supplementary winter feed were recorded as surviving a maximum of 7.7 and 8.7 years (female and male, respectively).76 The longest life span recorded for a female European rabbit in the wild is 7.6 years.65
Anatomy
Several books have been published on the anatomy of the rabbit. Unfortunately the two best atlases on rabbit anatomy are out of print. The more recent Colour Atlas of the Anatomy of Small Laboratory Animals: Rabbit, Guinea Pig has been republished three times since its first print in 1990.66 It is easily found in many libraries. The other, Atlas of Rabbit Anatomy (in French and English), is found only in selected university libraries owing to its limited print pressing in 1973.5
Skin and Hair
Rabbit skin is very delicate compared with that of other exotic pet species, dogs, and cats. Unless care is taken in clipping fur for surgery, it is easy to tear or rip the skin. Because rabbit hairs are so fine, electric hair clippers require the use of thin blades (size 50, 1/125 in. [0.2 mm], or 40, 1/100 in. [0.25 mm]), which are normally not used with cats and dogs. Hair growth in rabbits occurs in periodic, orderly waves originating on the head and ventrum and spreading dorsally and caudally.80 This is most noticeable when hair grows back after clipping for surgery. Moulting in adult rabbits usually occurs twice a year (spring and fall).
Scent Marking Glands
Rabbits are strongly territorial, and both sexes have three glands used in scent-marking behavior: the chin glands, which are specialized submandibular glands opening onto the underside of the chin; the anal glands; and a pair of pocket-like perineal glands called the inguinal glands that are relatively large and often harbor a normal yellowish-brown deposit. The size of the glands and degree of marking are androgen-dependent and related to the level of sexual activity. Males mark more frequently than do females, dominants of both sexes mark more frequently than subordinates do, and dominants mark most in the presence of subordinate rivals. Under natural conditions, both bucks and does on their own territory, surrounded by their own odor and that of their clan, win two-thirds of all aggressive encounters.57 When a rabbit becomes dominant, a new compound appears in the chin gland secretion, 2-phenoxyethanol.38 This compound is a fixative used in the perfume industry and slows the release rate of compounds in chin gland secretions, enabling the scent to persist in the environment and not dissipate. The dominant rabbit thus reigns over the olfactory environment, just as it does the physical environment.
Sense Organs and Nervous System
Eye
Prince has described the anatomy and physiology of the rabbit eye in great detail in the old but not outdated The Rabbit in Eye Research.68
The optic nerve is above the horizontal midline of the eye, and examination of the fundus involves looking upward into the eye. Retinal vessels spread out horizontally from the optic disk; also, the rabbit has a depression or physiologic cup in the optic disc, as does the dog. Rabbits do not have a tapetum lucidum.37
The nictitating membrane or third eyelid is prominent, found in the medial canthus of the eye and associated with deep (harderian) and superficial (nictitans) orbital glands. Behind it and separated from the deep part of the nictitating membrane cartilage is the harderian gland, which has a small white upper lobe and a large pink lower lobe. The lower lobe is known as the deep gland of the nictitating membrane. Both the deep and superficial glands of the third eyelid can potentially prolapse.42 The deep gland prolapses more frequently than the superficial gland.82 The clinical condition is similar to “cherry eye” in dogs.
The harderian gland is wrapped behind the eyeball and is phylogenetically and anatomically associated with the nictitating membrane. It is believed that the harderian gland has a significant function in social behavior. Generally horseshoe-shaped and situated deep within the orbit, the harderian gland consists of a smaller, almost white (upper) lobe connected to a large pink (lower) lobe by a narrow strand medial to the optic nerve. The single excretory duct opens at the base of the nictitating membrane, and the milky secretion is made up of a complex mixture of lipids, protein, and the pigment protoporphyrin, which provides lubrication for the edges of the eyelid.24 Epiphora in rabbits often presents as a white overflow of tears because of impaired drainage or obstruction of the nasolacrimal drainage system—the white tears are a normal secretion of the harderian gland.45 Other orbital glands include the lacrimal and accessory lacrimal glands, located caudodorsal and ventral to the orbit respectively.
In rabbits, the primary channel for return of venous blood from the head, including that from the eye, is the external jugular vein.67 In contrast, the primary drainage of the eye and head in humans is via the internal jugular vein. In other species, such as dogs, significant anastomoses exist between the branches of the internal and external jugular veins. In rabbits, such anastomoses are minor, and ligation or chronic catheterization of the external jugular vein results in swelling and protrusion of the eyeball for about 24 hours, after which its normal appearance returns.41 The same pattern of vascularity also applies to the arterial blood supply of rabbit’s eyes, but ligation of the external carotid artery results in ipsilateral ocular necrosis.
Rabbits, like rodents, have an extensive orbital venous plexus. Because of possible severe hemorrhage, enucleation of the eye is difficult compared with species such as dogs. Bilateral exophthalmos can occur in rabbits secondary to engorgement of the orbital venous sinuses (retrobulbar venous plexus). Rabbits with either compromised heart function or cranial thoracic masses,77 such as thymoma, thymic lymphosarcoma and thymic carcinoma, can present with marked “cranial vena cava” signs such as bilateral exophthalmos. Occasionally, bilateral exophthalmos can also be seen in rabbits exhibiting extreme fear.
The nasolacrimal drainage system provides a conduit for tears from the lacrimal lake to the nasal cavity. In rabbits, a single ventral lacrimal punctum (about 3-4 mm ventral to the lid margin), canaliculus, sac, duct, and nasal meatus form the drainage system for each eye. The diameter of the nasolacrimal duct is small and narrows in two places where it changes course. These two sites, the proximal maxillary bend and the apex of the main upper incisor, are important in the development of obstruction50 (see Chapter 37). Dacryocystitis and nasolacrimal duct obstruction (dacryostenosis) often cause epiphora, one of the most common ocular problems seen in rabbits (see chapter on ophthalmology).
Ear
The pinnae represent a large portion of the total body surface in rabbits, approximately 12% in New Zealand white rabbits.21 The pinnae are highly vascular and have the largest arteriovenous shunts in the body. At rest, the pinna of leporids is a thermoregulatory organ. Claude Bernard in 1852 implied that rabbits use their ears to lose heat6; at ambient temperatures above 30°C, there is a marked vasodilation of ears.69 The Australian population of the European rabbit has responded to its warmer environment by increasing the mean length of its pinna from 71 mm in England and France to 79 mm in Australia.62
Anatomists have hypothesized another role for the pinna of leporids as a part of a suspensory system for the greater portion of the head, absorbing shocks that might otherwise interfere with vision during high-speed locomotion.74 This is certainly the case with European hares. However, as the rabbit is semifossorial and does not rely on sustained fast running to avoid predation, as do hares, the significance of the ears in capital shock absorption in rabbits is less important.
In large-eared rabbits, the vessels of the ear margins (medial and caudal auricular veins) are easily accessible for venipuncture. However, in small-eared rabbits such as dwarf breeds, venipuncture of the ear veins can lead to vasculitis, vascular necrosis, and sloughing of the ear pinna. During anesthesia, the central auricular artery and auricular veins are good sites to obtain noninvasive measurements such as pulse oximeter blood oxygen saturation and laser-based systolic and diastolic arterial blood pressure.39
Muscles and Skeleton
Bones of rabbits are relatively delicate compared with their muscle mass. The skeleton represents only 7% to 8% of body weight in rabbits, whereas the skeletal muscle comprises more than 50% of the body weight.9,21 Fractures, especially of the tibia, are always a potential problem. In comparison, the skeleton of a cat constitutes 12% to 13% of body weight. However, the dry matter and percentage of calcium is higher in the bones of rabbits than those of cats.1
In countries where rabbit meat is commonly eaten, clients may present a headless carcass that they suspect to be that of a cat. The color of the muscles distinguishes the two: rabbit muscles are pale pink, whereas those of cats are deep red. There are also skeletal differences, most noticeably between the scapula and pelvis. The infraspinous fossa of the rabbit is sharply triangular, whereas that of the cat is more rounded; also, the suprahamate process of the acromion is truly hook-shaped in rabbits, whereas it is blunted in cats. Good drawings of these differences are found in Okerman’s Diseases of Domestic Rabbits.59
The acetabulum of rabbits is formed by the ilium, the ischium, and a small accessory bone, the os acetabuli, which excludes the pubis. In other animals, the acetabulum comprises the ilium, the ischium, and the pubis. The trochanteric fossa of the femur is a good site for intraosseous catheter placement and is easily located by palpating the prominent greater trochanter.10
Scientists have studied the architecture of skeletal muscle in different species to determine the force, velocity, and displacement properties of a muscle. The rabbit has a pelvic limb muscle mass of 13% (of total body mass), reflecting its need to evade predators by accelerating quickly and maintaining high-speed locomotion.83 Forelimb muscle mass accounts for 9% of total body mass, highlighting the predisposition toward hindlimb propulsion. The bulk of the fore- and hindlimb muscle mass is proximal; the distal limbs (e.g., carpi and tarsi) have little musculature; in the hindlimb, this adaptation decreases rotational inertia.46 Spinal musculature accounts for 9% of body mass—a substantial portion of the total locomotor muscle mass (~30%).83 Much of the back musculature extends the lumbar spine and increases the distance from the hindfoot to the rabbit’s center of mass. When normalized for body mass, the back muscles are much larger than those of the dog and horse. The powerful hindlegs and lumbar muscles mean that rabbits can kick violently. If they are not held securely when picked up, their kicking can result in a vertebral fracture (usually at the seventh lumbar vertebra) and damage to the spinal cord. Clinically, the large lumbar and proximal hindlimb muscles are the best sites for intramuscular injections.
The number of thoracic (T) and lumbar (L) vertebrae varies from 12T, 7L in 44%; 13T, 6L in 33%; and 13T, 7L in 23% of rabbits.33 The spinal cord ends within the second sacral vertebra (S2) in 79%, within the first sacral vertebra (S1) in 19%, and within the third sacral vertebra (S3) in 2%.33 This feature is unusual, as the spinal cord ends in other species (e.g., dog, horse) within the caudal lumbar region.
Digestive System
The definitive references on the anatomy and physiology of the digestive tract of rabbits are Cheeke’s Rabbit Feeding and Nutrition14 and the more recent The Nutrition of the Rabbit by de Blas and Wiseman.22 Most of the information described in this section is from these books unless otherwise noted.
Teeth
All the teeth of rabbits are classified elodont (continuously growing, with no anatomic “roots”) and hypsodont (long-crowned). The teeth can also be classified as aradicular. This leads to a dynamic state intrinsic to rabbit dentition; clinically, it serves to complicate dental disease. This type of dentition also allows for a concurrent increase in tooth size with growth of the animal.36
The rabbit dental formula is 2 (I2/1, C0/0, PM3/2, M3/3) = 28. It is useful clinically to divide rabbit dentition into two groups, the incisor teeth set and cheek teeth set (premolars and molars). Each cheek teeth arcade can be divided into quadrants (upper and lower on right and left). The rabbit mouth also features a relatively long diastema. The lower jaw is narrower than the upper jaw, a feature known as anisognathism. The maximum gape of the rabbit is only 20 to 25 degrees, compared with the rat’s 40 degrees.36 This, coupled with the long diastema, can make inspection of the oral cavity relatively difficult. Each tooth can be further divided into the clinical crown (region of tooth exposed above the gingival margin) and the reserve crown (region of tooth buried below the gingival margin). This reserve crown is often misnamed the root of the tooth. The growing portion at the tip of the reserve crown is the apex, which is open in the rabbit’s elodont hypsodont teeth (Fig. 12-3).
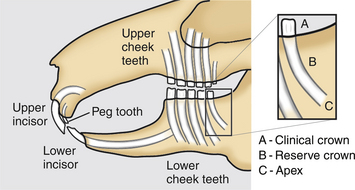
Incisors have a single pulp cavity. The cheek teeth have a single pulp chamber at the tooth apex that diverges into two toward the clinical crown. Enamel is present only on the labial (facing toward the lips) side of the strongly curved maxillary incisors, while the mildly curved mandibular incisors feature enamel on both the labial and lingual (facing toward the tongue) aspects of the teeth.20 Through normal wearing, this feature produces the characteristic chisel shape of the clinical crown tips (in occlusively normal incisor teeth). In contrast, the cheek teeth are longitudinally straighter and somewhat “folded” on the buccal (toward the cheek) side in cross section.36 This formation creates an increase in the proportion of enamel on the occlusal surface of the tooth (providing increasing grinding efficiency and helping to reduce the wear rate). The more heavily calcified alveolar bone surrounding each tooth socket is termed the lamina dura.
Normally occluded mandibular incisor tips lie just caudal to the main maxillary incisors, in the space between the primary maxillary incisors and the peg teeth (the smaller second set of upper incisor teeth situated just caudal to the main maxillary incisors).20 The tips of the mandibular incisors in occlusively normal rabbits make contact with the peg teeth.36 Rabbits periodically “grind” their teeth to help shape their incisor tips to their characteristic chisel shape.36
At rest, the opposing cheek teeth quadrants are normally in contact.36 This observation contrasts to what many authors have previously described, where they stated that at rest, the rabbit’s opposing cheek teeth quadrants do not normally contact each other. In fact, in the normal resting closed mouth gape, both the opposing incisors and cheek teeth are in contact with each other.36 During mastication, each lower cheek tooth occludes with two upper cheek teeth except for the first and last lower cheek teeth.36
The rate of tooth growth varies between the different sets and can be influenced by age, pregnancy, and diet.36 For example, the upper incisors grow more slowly than the lower incisors, at rates of 2 and 2.4 mm/week, respectively.72
Rabbits primarily use a vertical action to “cut” foliage with their incisors.36 Once typical leaf material is present in the mouth, it is then masticated largely in a horizontal or lateral plane by the cheek teeth. This is important to consider, as dietary factors have been shown to affect a rabbit’s normal chewing process.78 Food is ground by only one side of the cheek teeth at a time.36 Natural vegetation such as grass complements a “normal” horizontal chewing action of the cheek teeth, while harder and thicker food items (such as pellets, grains, or carrots) tend to encourage more vertical and less horizontal movements in mastication.78 This vertical action can lead to a reduction in tooth wear and also potentially increase forces on teeth in the vertical plane, producing increased pressure on the growing apex of the tooth.12 Further discussion of rabbit dental anatomy and physiology may be found in Chapter 32.
Small Intestine
The small intestine is shorter in rabbits than in other species, making up about 12% of the total volume of the gastrointestinal tract. The duodenum and jejunum have a relatively small lumen. Peyer’s patches are absent from the duodenum and the first half of the ileum. No correlation exists between the number of Peyer’s patches and intestinal length; surgeons or pathologists may see only three to nine Peyer’s patches in the first quarter of the jejunum and last quarter of the ileum.54,70 The terminal portion of the ileum ends with the rounded sacculus rotundus at the ileocecocolic junction. The sacculus rotundus has a minute honeycombed external appearance owing to the presence of a large number of lymph follicles; it is sometimes referred to as the ileocecal tonsil. The terminal portion of the ileum is a common site of foreign-body impaction due to luminal narrowing.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


