CHAPTER 15 Auditory System
Special Somatic Afferent System
ANATOMY
Receptor
The development of the bony and membranous labyrinths of the receptor in the auditory system is described with the development of the vestibular system in Chapter 12. Fig. 12-1 outlines the basic components; Fig. 15-1 outlines the basic components; shows their relationship to the other components of the ear. The cochlea is the coiled portion of the bony labyrinth in the petrosal portion of the temporal bone that contains perilymph. The degree of coiling varies with the species of animal. There are 3.5 turns in the dog cochlea, compared with 2.5 turns in the human cochlea. The portion of the bone that forms the center, or axis, of the coiled cochlea is the modiolus. It contains the cochlear division of the vestibulocochlear nerve. At the base of the modiolus, the cochlea is continuous with the vestibule, and there is an opening into the tympanic cavity that is called the cochlear window. A shelf of bone projects into the cochlea from the modiolus. This is the spiral lamina, which partially divides the cochlea into two portions and is absent at the apex, or most distal extent of the cochlea Fig. 15-2.
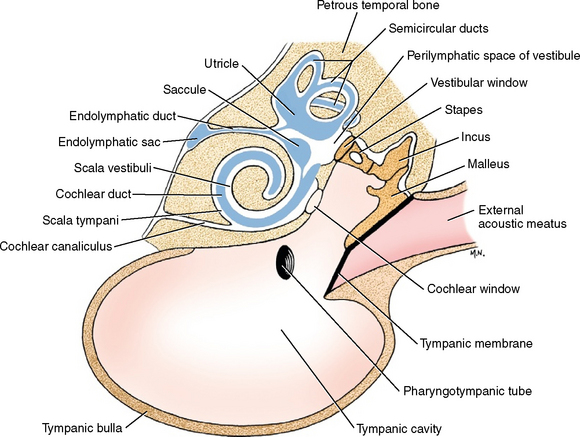
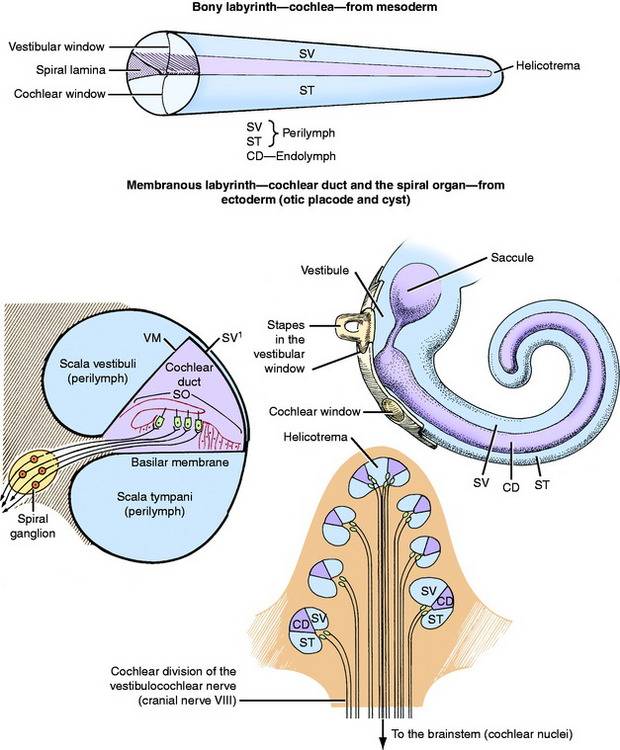
The cochlear duct is the coiled portion of the membranous labyrinth derived from the embryonic ectodermal otocyst. It is located inside the cochlea and contains endolymph. It is a tubular structure that, in transverse section, is triangular in shape. The apex of the triangle attaches medially to the spiral lamina, and the base of the triangle attaches laterally to the outer wall of the cochlea (see Fig. 15-2). This completes the partitioning of the cochlea into two portions, each filled with a continuous column of perilymph. The two sections are the scala vestibuli and the scala tympani. The scala vestibuli is situated dorsal to the cochlear duct; it communicates proximally with the vestibule and distally at the apex of the cochlear duct, with the scala tympani. The latter occurs because the cochlear duct does not reach the apex of the cochlea, leaving an opening between the two scali that is known as the helicotrema. The scala tympani is situated ventral to the cochlear duct. It communicates distally, at the helicotrema, with the scala vestibuli. Proximally, at the base of the coiled cochlea, the scala tympani terminates at the cochlear window, which is covered by a membrane that retains the perilymph within the bony labyrinth. On the other side of this membrane, at the cochlear window, is the air-filled tympanic cavity of the middle ear in the tympanic bulla. Perilymph fills the scala vestibuli and scala tympani (see Fig. 15-1). A small perilymphatic duct connects the scala tympani with the subarachnoid space medial to the petrosal portion of the temporal bone via the cochlear canaliculus. At the level of the origin of the cochlea from the vestibule, the cochlear duct communicates with the saccule by way of the ductus reuniens. Figs. 15-3 through 15-5 show the microscopic features of this anatomy.
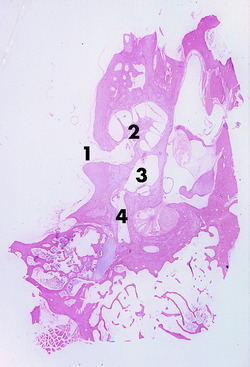
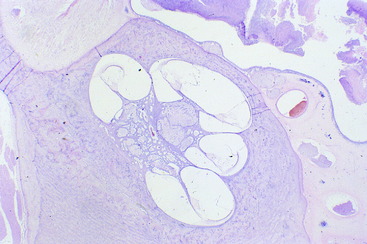
Figure 15-4 Higher magnification of the cochlea and cochlear duct coiled around the modiolus of a 1-month-old calf.
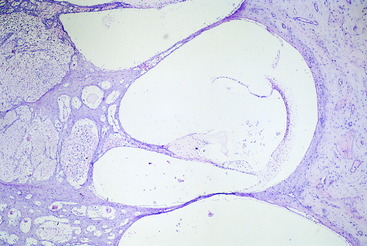
Figure 15-5 Higher magnification of one complete transverse section of the cochlea and cochlear duct of the calf in Fig. 15-4. The stria vascularis is on the right, with artifactitious separation from the wall of the cochlea.
The base of the triangular cochlear duct, when viewed in transverse section, is attached to the outer wall of the bony cochlea and consists of a stria vascularis and a spiral prominence (see Figs. 15-2 and 15-5). A thin layer of tissue called the vestibular membrane forms the dorsal border of the cochlear duct. This membrane is between the perilymph of the scala vestibuli and the endolymph of the cochlear duct. It extends from the stria vascularis dorsolaterally to the spiral lamina medially. The basilar membrane extends between the spiral lamina and the ventrolateral extent of the stria vascularis at the middle of the lateral wall of the cochlea (Fig. 15-6; see also Fig. 15-5). The basilar membrane is a highly organized layer of collagen fibers. The spiral organ (organ of Corti) is the specialized sensory neuroepithelium that is located on this basilar membrane.31 This spiral organ is composed of hair cells and several types of supporting cells. The hair cells have, on their luminal surfaces, modified microvilli known as stereocilia. The tips of these stereocilia are embedded in a proteinaceous membrane, the tectorial membrane, which covers the hair cells and is attached medially along the apex of the cochlear duct. The hair cells are associated with various types of supporting cells located on the basilar membrane. The dendritic zones of the neurons in the cochlear portion of the eighth cranial nerve are in synaptic relationship with the bases of the hair cells.29
Sound waves are transmitted from the air medium of the external ear canal to the solid medium of the tympanumand the chain of three ear ossicles, which extends to the vestibular window, and to the fluid medium of the perilymph in the scala vestibuli. Wave flow through the perilymph in the scala vestibuli is reflected to the basilar membrane by way of the vestibular membrane and the endolymph of the cochlear duct or by continuing through the helicotrema to the perilymph in the scala tympani. Movement of the highly organized basilar membrane causes the hair cells of the overlying spiral organ to move and their stereocilia, embedded in the tectorial membrane, to bend. This action causes an impulse to be generated in the cochlear neurons. The basilar membrane acts as a resonator, and different portions respond maximally to sound waves of specific frequencies. This function may be related to the length of the basilar membrane, which is longest at the base of the cochlear coil and progressively shortens through the coils to the apex. Low frequencies cause maximal vibration of the basilar membrane at the base of the cochlear duct. High frequencies affect the apical portion of the basilar membrane maximally. In dogs, the basilar membrane is sensitive to a frequency range of 2 to 47 Hertz, compared with a range of 1 to 19 Hertz in humans.
Cranial Nerve VIII: Vestibulocochlear Nerve, Cochlear Division
The neurons of the vestibulocochlear nerve are derived from placode ectoderm associated with the otocyst. The dendritic zone of the cochlear division of the eighth cranial nerve is in synaptic relationship with the base of the hair cells in the spiral organ. The axons course medially along the basilar membrane into the modiolus. The cell bodies of these bipolar neurons are located in the modiolus at the origin of the spiral lamina, where they form the spiral ganglion. The axons course though the center, or axis, of the modiolus to the internal acoustic meatus, where they are joined by the vestibular division of the eighth cranial nerve. Within the vestibulocochlear nerve, the axons of the cochlear division course to the region of the cerebellomedullary angle at the junction of the medulla and the pons, caudal to the transverse fibers of the pons. They terminate in telodendria on cell bodies in the dorsal and ventral cochlear nuclei that are located on the side of the medulla where the nerve enters. These nuclei bulge from the lateral side of the rostral medulla where they appear to be in the vestibulocochlear nerve (see Figs. 2-11, 2-12).
Brainstem Nuclei and Tracts
The axons of the cell bodies in the cochlear nuclei pass into the medulla by one of two pathways: ventrally through the trapezoid body or dorsally over the caudal cerebellar peduncle by way of the acoustic stria (see Figs. 2-12, 2-13). Numerous pathways involving a number of synapses are available in the brainstem to the auditory system for reflex activity and conscious perception (Fig. 15-7).
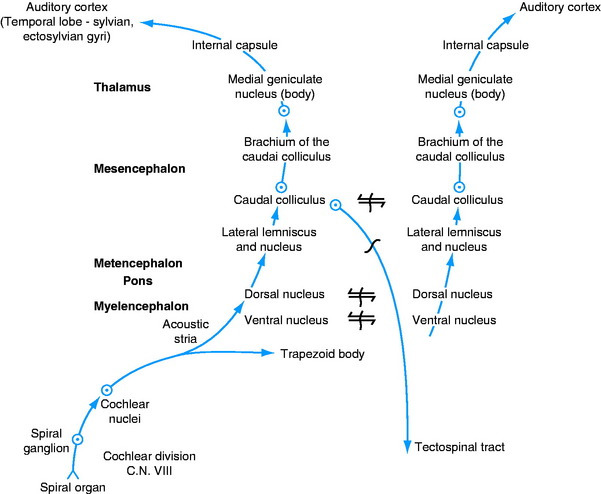
Reflexes are mediated directly, through the influence of cochlear nuclei neurons on the lower motor neurons (LMNs) in brainstem nuclei; or they are mediated indirectly by transmission through the neurons of the caudal colliculus or other auditory nuclei in the brainstem. Reflex regulation of the sound wave frequency occurs by way of the afferent neurons of the cochlear division of the eighth cranial nerve, the cochlear nuclei, and the efferent neurons of the motor nucleus of the trigeminal nerve in the pons that innervate the tensor tympani muscle and the efferent neurons in the facial nucleus in the medulla that innervate the stapedius muscle. These muscles control the degree of mobility of the ear ossicles.
Other neurons that belong to the auditory system and function in both reflex and conscious perception pathways include the dorsal and ventral nuclei of the trapezoid body, the nucleus of the lateral lemniscus, and the caudal colliculus.12 The neuronal cell bodies of the ventral nucleus of the trapezoid body are scattered without specific arrangement throughout the axons of the trapezoid body. The dorsal nucleus of the trapezoid body forms a distinctly encapsulated nucleus ventrally in the rostral medulla, dorsal to the trapezoid body and dorsolateral to the pyramid, from the level of the facial nucleus in the medulla to the motor nucleus of the trigeminal nerve in the pons (see Figs 2-11, 2-12). The lateral lemniscus is composed mostly of ascending axons in the auditory system as they course from the rostral medulla through the pons to the caudal mesencephalon on the lateral surface of the brainstem (see Fig 8 through 10).12 The lateral lemniscus contains axons from the cochlear nuclei or the various nuclei of the trapezoid body on the same side or the opposite side. The lateral lemniscus is formed medial to the middle cerebellar peduncle and is exposed on the lateral surface of the caudal mesencephalon rostral to the transverse fibers of the pons that form the middle cerebellar peduncle. Embedded along the ventromedial aspect of this lateral lemniscus is the nucleus of the lateral lemniscus (see Fig. 2-9). The lateral lemniscus terminates at the caudal colliculus.
The caudal colliculus consists of cell bodies of neurons and axonal processes organized in laminae (see Figs 2-8, 2-9).23 It functions as a reflex center for the auditory system. Efferent axons project to LMNs in the brainstem by way of the tectonuclear pathway and to LMNs in the cervical spinal cord through the tectospinal pathway in the ventral funiculus. The latter arises from the rostral colliculus. Efferent axons project to the thalamus in the brachium of the caudal colliculus located on the lateral side of the mesencephalon ventral to the rostral colliculus (see Figs 2-7, 2-8). They terminate in the medial geniculate nucleus (see Figs 2-5, 2-6).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


