Fig. 4.1
Localization of Culicoides surveillance sites in France, 2002–2009
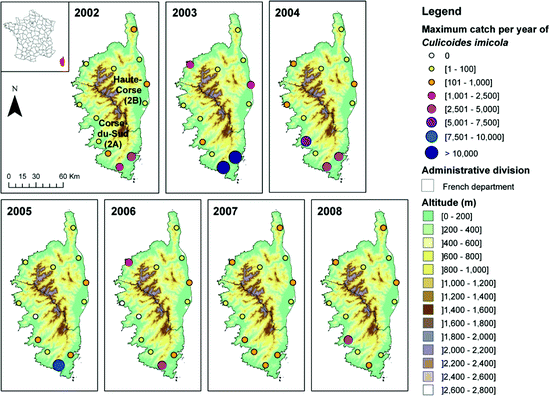
Fig. 4.2
Collection sites in Corsica with maximum catches of Culicoides imicola per year from 2002 to 2008
Moreover, from 2002, 19 sites along the Mediterranean coast of the French mainland (involving seven departments (administrative units): Pyrénées-Orientales, Aude, Hérault, Gard, Bouches-du-Rhône, Var and Alpes-Maritimes) (Fig. 4.1) were sampled, one night collection per site every month (Baldet et al. 2004). Additional traps were used in departments close to known established C. imicola populations in countries bordering France: in Pyrénées-Orientales and Alpes-Maritimes and in Pyrénées-Atlantiques in 2005 (Fig. 4.1). Indeed, in neighbouring countries, the closest populations of C. imicola are present in Spain, in Catalonia Peralda (Girona, Spain) (Sarto i Monteys et al. 2005) and in the Basque Country (Goldarazena et al. 2006), and in Italy (Torina et al. 2004). The Mediterranean littoral surveillance system is largely developed below in the section entitled “First Record of Culicoides imicola in Mainland France and Surveillance of Its Extension”.
In 2006, when BTV-8 emerged in northern Europe, France was initially marginally affected with only seven outbreaks along the Belgian border. In late September 2006, four cattle farms in the Ardennes department were thus sampled one night weekly to confirm the absence of C. imicola, describe Culicoides diversity and the end of their activity period (Baldet et al. 2008). Due to the large BTV spread in Belgium, the Netherlands and Germany in late 2006, the surveillance network was extended early 2007 to 15 farms along the northern border, with outdoor and indoor traps (Fig. 4.1). In late 2007, following the disease front evolution, the surveillance included 13 farms in the centre of France (Fig. 4.1). The aim of this network was (1) to determine the start and the end of the active period of Culicoides populations by trapping one night per week from March to April and from October to December and (2) to follow up Culicoides dynamics via monthly collections from May to September. At that time, the transmission of BTV by autochthonous Culicoides species occurred massively throughout the whole French territory (15,253 infected farms with BTV-8 in 2007, 23,959 infected farms with BTV-8 in 2008 and 4,339 infected farms with BTV serotype 1 in 2008). Indeed, BTV-1 was introduced in the Basque country from southern Spain and spread mainly in south-western France. The trapping frequency was standardized in 2008 for all traps of French mainland, with weekly collections at the start or end of Culicoides activity (from February to April and from October to December) and monthly collections the rest of the year. Trap localizations remained unchanged except in the Mediterranean littoral where the number of traps was reduced to 13 traps. Finally, from 2009 to date, in order to obtain information on the whole territory, the number of traps was increased to 160 distributed in the mainland and Corsica with 1–2 traps per department monitored at the same frequency as described just above (Balenghien et al. 2010, 2011).
In all surveillance catches, all Culicoides specimens were sorted by species, sex and physiological stages for females (nulliparous, parous or engorged). Subsampling following a standardized procedure was applied when abundance was high.
4.2 First Record of Culicoides imicola in Mainland France and Surveillance of Its Extension
Following the first record of C. imicola in August 2000 in Sardinia, Italy (Goffredo et al. 2001), and given the short distance between Sardinia and Corsica, France (12 km), French authorities mandated an entomological surveillance of the island early in October in order to establish whether Culicoides imicola was present (and established) on the island and thereafter assess the risk of BTV spreading to French mainland (Fig. 4.1). Early October 2000, 2 weeks before the first BTV outbreaks in Corsica, an entomological survey was conducted in nine farms during a week, with one night trapping using UV-light suction traps (Rieb trap) (Fig. 4.3).

Fig. 4.3
The two models of UV-light trap used during Culicoides surveillance in France. Rieb traps (picture from IPPTS) were used from 2000 to 2006 in Corsica, from 2002 to 2005 on the Mediterranean littoral, and in 2006 in the southwest France (a). OVI traps (picture from Vet Services, department Indre) were used in 2006 onwards in all trapping sites except Corsica where they were implemented in 2007 (b)
This survey recorded for the first time C. imicola in Corsica (October 2000) (Delécolle and de La Rocque 2002). It also highlighted that the species could be very abundant (with a single-night catch of 12,000 individuals at Porto-Vecchio in the south-east) and distributed in almost all the studied sites throughout the island, notably on the littoral (Fig. 4.2). Given the proximity of Corsica to the French mainland (180 km), the authorities dread the establishment of these populations into the French mainland. Therefore, an entomological survey was carried out with one night trapping in eight locations for three departments (Hérault, Gard and Bouches-du-Rhône) along the Mediterranean littoral in May 2001 and in the Var department in October 2011 (Fig. 4.1). A total of 419 Culicoides were trapped, and no C. imicola individuals were captured. In Corsica, in June 2001, 12 farms were prospected during a week confirming the presence of C. imicola in high numbers. In 2002, a wider surveillance programme was implemented in Corsica (12 sites) and in the French mainland (19 sites) (Figs. 4.1 and 4.2). This network enabled to trap the first C. imicola individuals on the mainland in 2003: one male the 22nd of May, in the Var department, and one nulliparous female the 24th of September in the Alpes-Maritimes department (Fig. 4.4), confirming that individuals could spread passively from Corsica. Additional nights of trapping conducted in and around both positive sites failed to confirm the presence of established populations.
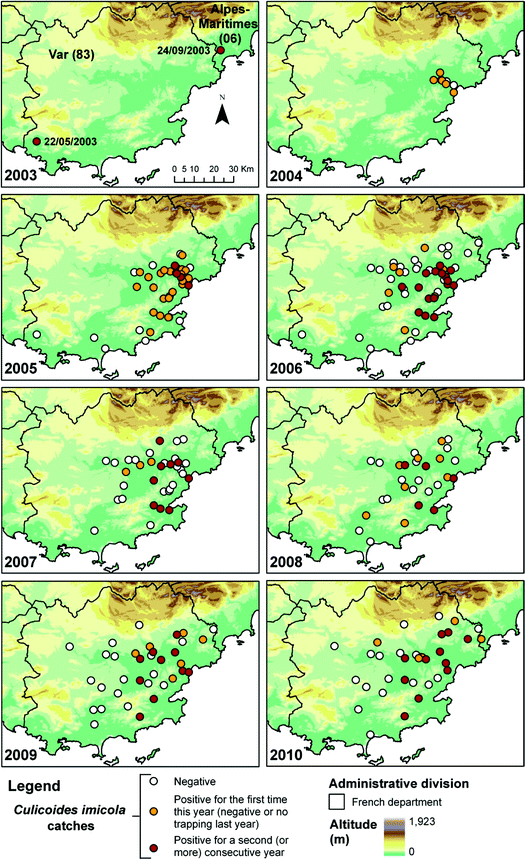
Fig. 4.4
Distribution area of Culicoides imicola in the Var department from 2003 to 2010
These catches coincided with a year of high abundance of C. imicola within Corsica and, particularly, in the north-east of the island in 2003 (Figs. 4.2 and 4.8). The next year, in June 2004, a gravid female was collected by the surveillance network in the valley of Argens (Var department), followed by one parous female late September, one nulliparous and two parous females early October. An expected critical zone of establishment was defined in the valley of Argens. Thus, additional trappings were carried out in October 2004 in this area (Table 4.1, Fig. 4.4).
Table 4.1
Summary of Culicoides catches in the Var department, 2004–2010
Year | 2004 | 2005 | 2006 | 2006 | 2007 | 2008 | 2009 | 2010 |
|---|---|---|---|---|---|---|---|---|
Sampling dates | 19-20/10 | 26/09-10/10 | 19-23/09 | 26-29/09 | 01-04/10 | 29/09-04/10 | 28/09-02/10 | 06-10/09 |
Number of sites | 16 | 42 | 51 | 45 | 34 | 29 | 35 | 34 |
Negative sites (0 C. imicola) [% total] | 6 [37.5] | 12 [28.6] | 40 [78.4] | 22 [48.9] | 22 [64.7] | 16 [55.2] | 19 [54.3] | 17 [50] |
Positive sites with 1–10 C. imicola [% total] | 7 [43.75] | 21 [50] | 8 [15.7] | 13 [28.9] | 6 [17.6] | 7 [24.2] | 11 [31.4] | 11 [32.3] |
Positive sites with 11–100 C. imicola [% total] | 2 [12.5] | 5 [11.9] | 2 [3.9] | 7 [15.5] | 2 [5.9] | 3 [10.3] | 2 [5.7] | 5 [14.7] |
Positive sites with >100 C. imicola [% total] | 1 [6.25] | 4 [9.5] | 1 [2] | 3 [6.7] | 4 [11.8] | 3 [10.3] | 3 [8.5] | 1 [2.9] |
Number of C. imicola | 240 | 4,636 | 257 | 8,935 | 948 | 1,348 | 546 | 1,497 |
Number of other Culicoides | 379 | 6,954 | 27,056 | 2,112 | 3,420 | 1,264 | 2,159 | 3,270 |
Total number of Culicoides | 619 | 11,590 | 27,313 | 11,047 | 4,368 | 2,612 | 2,705 | 4,770 |
Table 4.2
Maximal spread of Culicoides imicola over the years in the Var department
Year | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2004–2010 |
|---|---|---|---|---|---|---|---|
Number of traps positive for the first time | 25 | 7 | 2 | 4 | 4 | 6 | Total = 48 |
Maximal distance between a trap positive for the first time that year and its nearest positive trap of the preceding years (in km) | 30.66 | 11.60 | 5.52 | 18.89 | 10.44 | 9.84 | Max = 30.66 Mean = 14.49 |
This extensive trapping during 2 consecutive nights in 16 sites localized in the critical zone revealed that C. imicola was present in 10 sites, with a total of 240 C. imicola trapped out of 619 Culicoides. It thus confirmed the presence and lasting establishment of C. imicola on the French mainland (Fig. 4.4). Since 2004, the different C. imicola populations established in the valley of Argens have been surveyed each year by extensive trapping conducted during one night in several sites at the end of the summer to follow their geographical extension (Table 4.1). From 2003 to 2010, a gradual extension of the distribution area of C. imicola in the Var department was observed (Table 4.1, Fig. 4.4).
In 2007, out of the 34 studied sites, 12 were positive for C. imicola, with four sites separated by a few kilometers collecting 90% of the total number of C. imicola individuals (Fig. 4.4). One isolated population was detected at Grimaud, south of the Argens valley. In 2008, C. imicola populations remained restricted to some areas: the valley of Argens, the coastal plains of Fréjus and St Tropez golf, and some sites such as Cannet-des-Maures, Grimaud, Bormes-les-Mimosas and Pierrefeu-du-Var. In 2009, despite important floods in June in the known distribution area of C. imicola, new positive sites were recorded eastward (Mandelieu-la Napoule, département Alpes-Maritimes, Callian, département Var), highlighting an eastward expansion. Up to now, the distribution area of C. imicola seemed to be limited by physical barriers such as the massifs of Estérel and Tanneron (600 m) to the east, planes of Canjuers (1,000 m) and la Colle du Rouet (500 m), slowing down its progression to the east (Cannes), south-east (Hyères) and north (Fayence) of the Var department (Figs. 4.5 and 4.6).
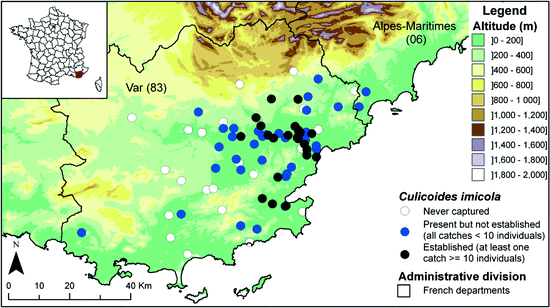
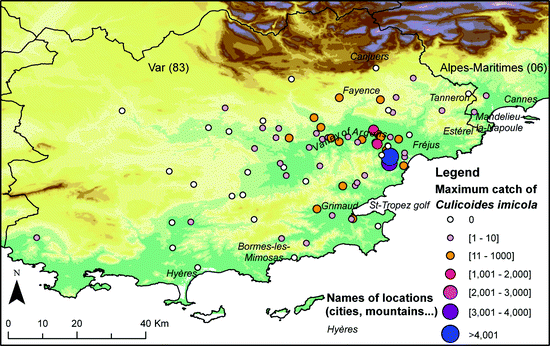
Fig. 4.5
Distribution area of Culicoides imicola in the Var department based on catches from 2003 to 2010
Fig. 4.6
Maximum catch of Culicoides imicola in the Var department from 2003 to 2010
To estimate the spread of C. imicola over the years, the distance between each trap site which was positive in a given year and the closest nearest neighbouring site which was found positive at least once during the preceding years was computed. The maximal observed spread of C. imicola per year corresponds to the maximal distance between two nearest neighbours for that year. This relies on the hypothesis that C. imicola has spread between sites following the shortest possible path. The sites positive in 2003 were not included as they did not lead to the establishment of C. imicola populations the following year.
Over the 2004–2010 period, the average spread was 14.5 km/year. The maximal observed spread was 30.7 km/year and occurred in 2005. That year, the spread may have been overestimated as in 2004, the trapping campaign was not large enough to encompass a ring of negative trapping sites around positive sites, indicating that C. imicola may have spread further than what was observed in 2004. The speed of colonization of this species over land seems to be limited by topography and vegetation cover (Tran et al. 2007) (Figs. 4.5 and 4.6).
The potential extension of C. imicola from Spain to the Pyrénées-Orientales department was also surveyed starting 2009 since this species is permanently established in the Spanish bordering area (Catalonia). In October 2008, three erratic individuals were collected by the surveillance network at the lowest point along the France-Spain border in the Pyrénées-Orientales department. To check whether C. imicola was absent from this region, additional trappings were carried out in October 2009 but did not collect any Culicoides because of heavy rains and wind. The following year, in October 2010, 11 sites were sampled for a week with one night trapping, and one site was positive with one parous female. The site was localized closed to the highway, suggesting that erratic individuals may passively be transported through the Pertus pass. Surveillance in this area is maintained, as this area is identified as risk of C. imicola establishment despite the efficient natural barrier of the Pyrenees Mountains and the North to South dominant winds.
4.3 Species Diversity
Through these surveys, the fauna list of Culicoides of France was updated. For most of the collections, traps were set up close to the hosts of interest (cattle and/or sheep); therefore, the checklist is probably not exhaustive and only related to the biting midge species related to domestic ruminants and therefore with a veterinary interest. Establishment of faunistic inventories is important to list the biodiversity, to analyse the possible spread of species into new territories and to analyse species communities in relation to the environment. Species diversity varies depending on the environment, reflecting differences in soil composition and meteorological conditions, and locally different breeding practices. When assessing the diversity of species to record rare or locally distributed species, it is important to keep in mind that collecting efforts often vary over time and that the type of trap used may bias the diversity of species sampled; for example, light traps are not appropriate to collect diurnal species.
Before bluetongue emergence in France, Delécolle (1985) recorded 65 species over the mainland territory with 45 recognized species in the Northeast region. A first checklist was published by Kremer et al. (1971) for Corsica recording 11 Culicoides species. However, the first bluetongue outbreaks in 2000, the first record of C. imicola on the island (Delécolle and de La Rocque 2002) and the resulting entomological surveillance increased the number of species records. Zientara et al. (2000) reported 21 species, and Delécolle and de La Rocque (2002) later recognized 37 species with 7 new species for Corsica. In 2005, Delécolle et al. (2005) updated the list of Culicoides in Corsica to 58 species with the first description of C. riebi. Today, the species list for French biting midges includes 83 species for mainland and 61 species for Corsica with recently three new records for mainland France: C. manchuriensis, C. abchazicus and C. saevus (Table 4.3). This list provides all the species collected through the surveillance networks plus some species recorded by experts but rarely collected (Delécolle and de La Rocque 2002; Delécolle et al. 2005). Whereas C. imicola can be dominant in Corsica, the common species collected in mainland France are the two sibling species of the Obsoletus Complex: C. obsoletus and C. scoticus usually referred as C. obsoletus/C. scoticus because females are morphologically indistinguishable, C. chiopterus, C. dewulfi, C. pulicaris, C. newsteadi and C. punctatus (Baldet et al. 2008; Balenghien et al. 2010, 2011) with some species locally being widely collected, as C. brunnicans in western France (Viennet et al. 2011). Culicoides chiopterus and C. dewulfi are rarely collected in Corsica.
Table 4.3
Updated list of Culicoides species recorded in France, and their distribution areas
Species name | Synonyms following the world biting midge catalogue | Distribution | |||
|---|---|---|---|---|---|
Mainland France | |||||
Atlantic coast | centre and northeast | Mediteranean littoral | Corsica | ||
C. abchazicus Dzhafaraov, 1964 | x (SR) | ||||
C. achrayi Kettle and Lawson, 1955 | x | x | x | x | |
C. alazanicus Dzhafarov, 1961 | C. musilator Kremer and Callot, 1961 | x | x | x | x |
C. albicans (Winnertz), 1852 | x | ||||
C. albihalteratus Goetghebuer, 1935a | x (SR) | x (SR) | |||
C. begueti Clastrier, 1957 | x | x (SR) | x | x | |
C. brunnicans Edwards, 1939 | x | x | x | x | |
C. cameroni Campbell and Pelham-Clinton, 1960 | x | x (SR) | |||
C. cataneii Clastrier, 1957 | x | x | x | x | |
C. caucoliberensis Callot, Kremer, Rioux and Descous, 1967 | X (SR) | ||||
C. chiopterus (Meigen), 1830 | C. amoenus (Winnertz), 1852; C. similis (Goetghebuer), 1927; C. dobyi Callot and Kremer, 1969 | x | x | x | x |
C. circumscriptus Kieffer, 1918 | C. nadayanus Kieffer, 1918 ; C. edwardsi Goetghebuer, 1921 ; C. algarum Kieffer, 1924 ; C. salicola Kieffer, 1924 ; C. pictidorsum Kieffer, 1924 ; C. albonotatus Vimmer, 1932 ; C. albosignatus Vimmer, 1932 ; C. polymaculatus Vimmer, 1932 ; C. pulcher Zilahi–Sebess, 1934 ; C. kirovabadicus Dzhafarov 1964 ; C. matsuensis Lien, Weng and Lin, 1996 ; C. meridionalis Xue, Liu and Yu, 2003 | x | x | x | x |
C. clastrieri Callot, Kremer and Déduit, 1962 | x | x | x (SR) | x | |
C. clintoni Boorman, 1984 | x | ||||
C. comosioculatus Tokunaga, 1956 | C. chaetophthalmus Amosova, 1957; C. caucasicus Sergejev, 1959; C. setosus Gutsevich, 1960 | x | |||
C. corsicus Kremer, Leberre and Beaucournu-Saguez, 1971 | x | ||||
C. dzhafarovi Remm, 1967 | C. dzhafarovi Callot, Kremer, Molet and Bach, 1968 | x | |||
C. deltus Edwards, 1939a | C. lupicaris Downes and Kettle, 1952 | x | x | x | x |
C. derisor Callot and Kremer, 1965 | x | x (SR) | |||
C. dewulfi Goetghebuer, 1936 | C. pseudochiopterus Downes and Kettle, 1952 | x | x | x | x |
C. duddingstoni Kettle and Lawson, 1955 | x | x | x | x | |
C. fagineus Edwards, 1939 | x | x | x | x | |
C. fascipennis (Staeger), 1839 | C. distictus Kieffer, 1916; C. albonotatus Kieffer, 1918; C. turficola Kieffer, 1925 | x | x | x | x |
C. festivipennis Kieffer, 1914 | C. odibilis Austen, 1921; C. winnertzi Edwards, 1926 | x | x | x | x |
C. flavipulicaris Dzhafarov, 1964 | x | x | x | x | |
C. furcillatus Callot, Kremer and Paradis, 1962 | x | x | x
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
 Get Clinical Tree app for offline access
Get Clinical Tree app for offline access

| ||